Compitistudiando questa sezione:
- Questa sezione aiuta a comprendere le vie dell'evoluzione del mondo organico, graduale complicazione non solo strutturale, ma anche meccanismi fisiologici e biochimici, che hanno portato alla nascita dei moderni cordati cranici, compresi i primati e l'uomo;
- nel processo di confronto è possibile verificare chiaramente la condizionalità fisiologica dell'apparizione o della complicazione di determinate strutture, nonché la loro scomparsa, ovvero è possibile comprendere meglio la struttura del corpo umano;
- aiuta a comprendere l'embriologia umana, perché secondo la legge biogenetica fondamentale formulata nel 1866 da Haeckel e Müller: l'ontogenesi è una breve e rapida ripetizione della filogenesi.
Pertanto, nell'embriogenesi, gli esseri umani sono caratterizzati dalla comparsa di strutture, funzioni e reazioni biochimiche che erano presenti nei loro antenati cordati. Tali ripetizioni sono chiamate riepiloghi. Nell'ontogenesi umana di solito non compaiono le strutture dei pesci adulti, degli anfibi e dei rettili, ma solo le loro larve. Inoltre, le prime fasi dello sviluppo ricapitolano in modo più completo di quelle successive.
Ma lo sviluppo evolutivo non è una semplice ripetizione delle caratteristiche strutturali degli antenati, ma le presuppone cambiamenti durante la formazione di organi e sistemi . Questo:
- cenogenesi - adattamenti dell'embrione a un ambiente diverso, che si perdono nelle forme adulte, ad esempio le branchie in un girino; organi provvisori degli amnioti;
- filembriogenesi - cambiamenti evolutivi che hanno significato adattivo:
UN) anabolica (estensione) - aggiunta di fasi aggiuntive, ad esempio il diaframma muscolare dei mammiferi, le piume degli uccelli;
B) deviazione - deviazione dal programma nelle fasi intermedie della morfogenesi, ad esempio la formazione di ossicini uditivi, la comparsa di una struttura alveolare dei polmoni anziché cellulare; in questi casi la legge biogenetica è parzialmente soddisfatta;
V) arcallassi - deviazione dell'ontogenesi all'inizio, quando la legge biogenetica non è soddisfatta, ad esempio, la formazione dei capelli inizia con un ispessimento dell'epidermide, che è immersa nel corion;
G) eterocronia - cambiamento nel tempo di formazione della struttura, ad esempio, formazione precoce dell'amnio placentare; E eterotopia - cambiando la posizione della struttura, ad esempio, la formazione del cingolo scapolare umano a livello delle vertebre cervicali III-IV, che poi si sposta al livello delle vertebre toraciche I-II.
Con i disturbi dell'embriogenesi, gli adulti possono sviluppare segni che erano presenti nei loro lontani antenati: atavismi. Se riducono la vitalità, vengono chiamati atavico(o ancestrale da anĉetre) vizi. Gli atavismi sorgono a causa di:
Sottosviluppo degli organi (ipoplasia del diaframma, palatoschisi - “palatoschisi”, ecc.);
Persistenza (conservazione) delle strutture embrionali (dotto pervio, fistole laterali del collo, fistole ombelicali, ecc.);
Disturbi nel movimento degli organi nell'ontogenesi (localizzazione pelvica degli organi, criptorchidismo, ecc.).
Consideriamo localizzazione dei cordati nel mondo animale e loro classificazione.
Il phylum Chordata unisce deuterostomi multicellulari di diversi livelli di organizzazione. Inoltre, assolutamente tutti i cordati, almeno in uno stadio di sviluppo, hanno tre caratteristiche che li distinguono dagli altri animali:
- un complesso assiale di organi, che si basa sullo scheletro interno - la notocorda, sopra - il tubo neurale, sotto - il tubo digestivo;
- la faringe è penetrata da fessure faringee, che nei cordati superiori sono sostenute da archi viscerali;
- sezione della coda
Inoltre, si può notare la presenza del sistema venoso della vena porta (il sangue dal tratto gastrointestinale passa attraverso il fegato) e la struttura del tegumento dell'epidermide e del derma, ma il tegumento degli echinodermi ha una struttura simile.
Il phylum Chordata comprende quattro sottophyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Cordati larvali (Urochordata) - ascidie;
- III. Senza cranio (Acrania) - lancette;
- IV. Vertebrati (Vertebrata), che comprendono diverse classi:
Gruppo Anamnia (vertebrati inferiori)
- Ciclostomi (Cyclostomata) - lamprede, missine - non hanno mascelle;
- superclasse Pesci:
UN. classe Pesci cartilaginei e corazzati (Chondrichthyes) - squali, razze, chimere;
B. classe Pesci ossei (Osteitti) - altri pesci;
- Anfibi (Anfibi):
UN. Animali dalla coda: tritoni, salamandre;
B. Animali senza coda: rane, rospi, raganelle;
V. Quelli senza gambe sono ceciliani;
Gruppo Amniota (vertebrati superiori)
- Rettili (Rettili):
UN. Squamoso: lucertole, serpenti;
B. tartarughe;
V. coccodrilli;
g.Beakheads - hatteria;
- Uccelli (Aves) - un ramo laterale dell'evoluzione, hanno molti adattamenti specifici
UN. Ratiti: struzzi;
B. Pinguini;
V. Chiglie - altri uccelli;
- Mammiferi (Mammalia):
UN. Oviparo: ornitorinco, echidna;
B. Marsupiali: canguri;
V. Placentati: altri animali e umani.
FILOGENESI DELL'INTEGRAZIONE ESTERNA
Le principali funzioni del tegumento sono la delimitazione e la protezione di un organismo multicellulare. Il tegumento dei vertebrati acquisisce numerose funzioni aggiuntive. Nei cordati il tegumento è costituito da:
Epidermide che si sviluppa dall'ectoderma;
Derma che si sviluppa dal mesoderma.
Le direzioni di evoluzione sono le seguenti:
L'epidermide monostrato (lancetta) diventa multistrato (nei vertebrati), gli strati si differenziano e si cheratiniscono all'esterno; compaiono i derivati dell'epidermide: scaglie cornee, artigli, peli, ecc.
Il tessuto connettivo sottile e gelatinoso del derma si ispessisce e acquista forza grazie all'aumento del numero di fibre.
L'epidermide e il derma hanno numerosi derivati: cromatofori, scaglie, scudi cornei, conchiglie, placche, becco, artigli, piume, peli, corna, zoccoli. Tutti i cordati hanno ghiandole. Negli animali inferiori si tratta di ghiandole mucose unicellulari, a partire dai pesci compaiono ghiandole velenose, poi ghiandole sebacee (negli uccelli), nei mammiferi si aggiungono le ghiandole mammarie e sudoripare.
Una persona può avere i seguenti disturbi dell'ontogenesi del tegumento: albinismo(completo) e vitiligine depigmentazione cutanea (incompleta), alopecia(assenza) e ipertricosi(crescita eccessiva) di capelli, ipercheratosi E pachionichia- ispessimento dell'epidermide e delle unghie, rispettivamente, politelia E polimastia- un aumento del numero di capezzoli e ghiandole mammarie, rispettivamente.
FILOGENESI SCHELETRICA
I cordati inferiori vivono nell'acqua, mentre i cordati superiori vivono principalmente sulla terra. Il passaggio da un ambiente acquatico più denso a un ambiente aereo ha portato a una significativa complicazione dello scheletro e dei muscoli, in particolare, lo scheletro degli arti e le loro cinture sono diventati più complessi e il numero di muscoli specializzati che servono gli arti è aumentato. Le principali funzioni dello scheletro: supporto, base per l'attaccamento muscolare, partecipazione al metabolismo minerale.
Lo scheletro assile si basa sulla notocorda, quindi le vertebre si sono formate attorno alla notocorda, si sono differenziate, il numero di parti della colonna vertebrale è aumentato, nell'uomo, a causa della postura eretta, si sono formate curve - 2 cifosi e 2 lordosi, e i resti di la notocorda era conservata solo nelle sezioni centrali dei dischi intervertebrali.
Così, nel pesce Ci sono due sezioni della colonna vertebrale:
Tronco (vertebre del tronco articolate con le costole);
Coda;
negli anfibi :
Vengono aggiunte le sezioni cervicale e sacrale (1 vertebra ciascuna);
nei rettili :
Cervicale (8 vertebre, I - atlante, II - asse);
Toracico (articolato con le costole, alcune delle quali articolate con lo sterno davanti);
Lombare, sacrale (contengono un numero maggiore di vertebre);
Coda;
negli uccelli :
Cervicale (fino a 25 vertebre);
Petto;
L'osso sacro lombare e parte delle vertebre caudali sono saldamente collegati, formando un potente sacro;
nei mammiferi :
Cervicale (7 vertebre);
Toracico (9-24 vertebre);
Lombare (3-9 vertebre);
Sacrale (le vertebre si fondono insieme);
Caudale (coccigeo).
Nell'uomo esistono diversi difetti nella formazione del canale spinale, attraverso il quale può emergere la spina bifida, così come la formazione di curve patologiche, comprese quelle laterali (scoliosi), e la regione caudale può essere preservata.
Lo scheletro della testa è diviso in cranio cerebrale e viscerale. Nel processo di evoluzione, si è verificata una diminuzione del numero di ossa del cranio cerebrale, la sostituzione delle parti cartilaginee con ossa e una diminuzione del numero di elementi mobili. Anche il numero degli archi branchiali dello scheletro viscerale è progressivamente diminuito. Il primo e il secondo arco branchiale sono cambiati e hanno dato origine alle mascelle, alle ossa del cranio facciale, alle ossa uditive e alla cartilagine della laringe.
Nei pesci, l'arco I (mascellare, costituito dal palatoquadrato e dalle cartilagini di Meckel) è direttamente collegato all'arco II (costituito da cartilagine iomandibolare e ioide). Questo tipo di connessione tra il cervello e il cranio facciale si chiama iostilo(poiché il II arco si collega alla cartilagine iomandibolare). A partire dagli anfibi il primo arco, con la sua cartilagine palatoquadrata, si fonde con la base del cranio. Questo tipo di connessione si chiama stile automobilistico. La colonna ossea uditiva è formata dalla cartilagine iomandibolare, dalla quale successivamente si forma la staffa. Con disturbi ontogenetici, una persona può avere solo una di queste ossa uditive (come negli anfibi e nei rettili) invece di tre.
Consideriamo la struttura dell'organo uditivo in varie classi di vertebrati. U pescare Esiste solo l'orecchio interno, che comprende il labirinto membranoso con otoliti, canali semicircolari e il rudimento cocleare.
U anfibi C'è un orecchio interno della stessa struttura e un orecchio medio con un osso uditivo e un timpano.
U rettili l'orecchio interno è costituito dalla coclea e dai canali semicircolari, l'orecchio medio è lo stesso degli anfibi, c'è un rudimento del canale uditivo esterno.
U uccelli il canale uditivo esterno è ben formato.
U mammiferi l'orecchio interno comprende la coclea 2,5 giri, canali semicircolari, sacculo, utricolo; orecchio medio - la cavità timpanica in cui si trovano le ossa uditive - la staffa (un residuo della cartilagine iomandibolare), l'incudine (un residuo della cartilagine palato quadrata), il martello (un residuo della cartilagine di Meckel), c'è un timpano membrana, quella esterna comprende il canale uditivo esterno e il padiglione auricolare.
Pertanto, nello scheletro viscerale, nel processo di evoluzione, le mascelle cartilaginee furono sostituite da quelle ossee, cambiò il tipo di attacco delle mascelle e cambiò la funzione degli elementi degli archi viscerali.
Nello scheletro degli arti si formarono cinture con uno scheletro di arti liberi fissato in modo mobile; si formò un arto a cinque dita di tipo terrestre; il numero delle ossa nelle sezioni distali è diminuito; le parti prossimali degli arti furono allungate e le parti distali accorciate.
Una persona può avere i seguenti difetti: polidattilia E polifalange(aumento del numero delle dita e delle falangi, rispettivamente); spostando la cintura degli arti superiori al livello delle I-II vertebre toraciche - La malattia di Sprengel; anomalie congenite delle costole e deformità del torace - acqua ad imbuto, carenato e così via.
FILOGENESI DEI MUSCOLI
I muscoli svolgono la funzione di movimento e si dividono in:
Somatico, che si sviluppa dai miotomi ed è innervato dai nervi spinali (striati);
Viscerale: si sviluppa da altre parti del mesoderma, innervato da nervi autonomi, può essere striato e liscio.
Nei cordati inferiori, i muscoli viscerali si trovano nella regione degli archi branchiali e gradualmente da esso si formano i muscoli della masticazione, i muscoli della faringe, quindi i muscoli del collo e del cingolo scapolare, nonché i muscoli facciali . I muscoli del collo e del viso hanno un'innervazione complessa, associata alle fasi della filogenesi. I muscoli somatici nei cordati inferiori si trovano sotto forma di miomeri, quindi si formano i muscoli degli arti, la forma, le dimensioni e le funzioni dei muscoli diventano più diverse, la loro innervazione aumenta, specialmente gli arti liberi nei primati. I muscoli del tronco si riducono nel processo di evoluzione.
Compitistudiando questa sezione:
- Questa sezione aiuta a comprendere le vie dell'evoluzione del mondo organico, graduale complicazione non solo strutturale, ma anche meccanismi fisiologici e biochimici, che hanno portato alla nascita dei moderni cordati cranici, compresi i primati e l'uomo;
- nel processo di confronto è possibile verificare chiaramente la condizionalità fisiologica dell'apparizione o della complicazione di determinate strutture, nonché la loro scomparsa, ovvero è possibile comprendere meglio la struttura del corpo umano;
- aiuta a comprendere l'embriologia umana, perché secondo la legge biogenetica fondamentale formulata nel 1866 da Haeckel e Müller: l'ontogenesi è una breve e rapida ripetizione della filogenesi.
Pertanto, nell'embriogenesi, gli esseri umani sono caratterizzati dalla comparsa di strutture, funzioni e reazioni biochimiche che erano presenti nei loro antenati cordati. Tali ripetizioni sono chiamate riepiloghi. Nell'ontogenesi umana di solito non compaiono le strutture dei pesci adulti, degli anfibi e dei rettili, ma solo le loro larve. Inoltre, le prime fasi dello sviluppo ricapitolano in modo più completo di quelle successive.
Ma lo sviluppo evolutivo non è una semplice ripetizione delle caratteristiche strutturali degli antenati, ma le presuppone cambiamenti durante la formazione di organi e sistemi . Questo:
- cenogenesi - adattamenti dell'embrione a un ambiente diverso, che si perdono nelle forme adulte, ad esempio le branchie in un girino; organi provvisori degli amnioti;
- filembriogenesi - cambiamenti evolutivi che hanno significato adattivo:
UN) anabolica (estensione) - aggiunta di fasi aggiuntive, ad esempio il diaframma muscolare dei mammiferi, le piume degli uccelli;
B) deviazione - deviazione dal programma nelle fasi intermedie della morfogenesi, ad esempio la formazione di ossicini uditivi, la comparsa di una struttura alveolare dei polmoni anziché cellulare; in questi casi la legge biogenetica è parzialmente soddisfatta;
V) arcallassi - deviazione dell'ontogenesi all'inizio, quando la legge biogenetica non è soddisfatta, ad esempio, la formazione dei capelli inizia con un ispessimento dell'epidermide, che è immersa nel corion;
G) eterocronia - cambiamento nel tempo di formazione della struttura, ad esempio, formazione precoce dell'amnio placentare; E eterotopia - cambiando la posizione della struttura, ad esempio, la formazione del cingolo scapolare umano a livello delle vertebre cervicali III-IV, che poi si sposta al livello delle vertebre toraciche I-II.
Con i disturbi dell'embriogenesi, gli adulti possono sviluppare segni che erano presenti nei loro lontani antenati: atavismi. Se riducono la vitalità, vengono chiamati atavico(o ancestrale da anĉetre) vizi. Gli atavismi sorgono a causa di:
Sottosviluppo degli organi (ipoplasia del diaframma, palatoschisi - “palatoschisi”, ecc.);
Persistenza (conservazione) delle strutture embrionali (dotto pervio, fistole laterali del collo, fistole ombelicali, ecc.);
Disturbi nel movimento degli organi nell'ontogenesi (localizzazione pelvica degli organi, criptorchidismo, ecc.).
Consideriamo localizzazione dei cordati nel mondo animale e loro classificazione.
Il phylum Chordata unisce deuterostomi multicellulari di diversi livelli di organizzazione. Inoltre, assolutamente tutti i cordati, almeno in uno stadio di sviluppo, hanno tre caratteristiche che li distinguono dagli altri animali:
- un complesso assiale di organi, che si basa sullo scheletro interno - la notocorda, sopra - il tubo neurale, sotto - il tubo digestivo;
- la faringe è penetrata da fessure faringee, che nei cordati superiori sono sostenute da archi viscerali;
- sezione della coda
Inoltre, si può notare la presenza del sistema venoso della vena porta (il sangue dal tratto gastrointestinale passa attraverso il fegato) e la struttura del tegumento dell'epidermide e del derma, ma il tegumento degli echinodermi ha una struttura simile.
Il phylum Chordata comprende quattro sottophyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Cordati larvali (Urochordata) - ascidie;
- III. Senza cranio (Acrania) - lancette;
- IV. Vertebrati (Vertebrata), che comprendono diverse classi:
Gruppo Anamnia (vertebrati inferiori)
- Ciclostomi (Cyclostomata) - lamprede, missine - non hanno mascelle;
- superclasse Pesci:
UN. classe Pesci cartilaginei e corazzati (Chondrichthyes) - squali, razze, chimere;
B. classe Pesci ossei (Osteitti) - altri pesci;
- Anfibi (Anfibi):
UN. Animali dalla coda: tritoni, salamandre;
B. Animali senza coda: rane, rospi, raganelle;
V. Quelli senza gambe sono ceciliani;
Gruppo Amniota (vertebrati superiori)
- Rettili (Rettili):
UN. Squamoso: lucertole, serpenti;
B. tartarughe;
V. coccodrilli;
g.Beakheads - hatteria;
- Uccelli (Aves) - un ramo laterale dell'evoluzione, hanno molti adattamenti specifici
UN. Ratiti: struzzi;
B. Pinguini;
V. Chiglie - altri uccelli;
- Mammiferi (Mammalia):
UN. Oviparo: ornitorinco, echidna;
B. Marsupiali: canguri;
V. Placentati: altri animali e umani.
FILOGENESI DELL'INTEGRAZIONE ESTERNA
Le principali funzioni del tegumento sono la delimitazione e la protezione di un organismo multicellulare. Il tegumento dei vertebrati acquisisce numerose funzioni aggiuntive. Nei cordati il tegumento è costituito da:
Epidermide che si sviluppa dall'ectoderma;
Derma che si sviluppa dal mesoderma.
Le direzioni di evoluzione sono le seguenti:
L'epidermide monostrato (lancetta) diventa multistrato (nei vertebrati), gli strati si differenziano e si cheratiniscono all'esterno; compaiono i derivati dell'epidermide: scaglie cornee, artigli, peli, ecc.
Il tessuto connettivo sottile e gelatinoso del derma si ispessisce e acquista forza grazie all'aumento del numero di fibre.
L'epidermide e il derma hanno numerosi derivati: cromatofori, scaglie, scudi cornei, conchiglie, placche, becco, artigli, piume, peli, corna, zoccoli. Tutti i cordati hanno ghiandole. Negli animali inferiori si tratta di ghiandole mucose unicellulari, a partire dai pesci compaiono ghiandole velenose, poi ghiandole sebacee (negli uccelli), nei mammiferi si aggiungono le ghiandole mammarie e sudoripare.
Una persona può avere i seguenti disturbi dell'ontogenesi del tegumento: albinismo(completo) e vitiligine depigmentazione cutanea (incompleta), alopecia(assenza) e ipertricosi(crescita eccessiva) di capelli, ipercheratosi E pachionichia- ispessimento dell'epidermide e delle unghie, rispettivamente, politelia E polimastia- un aumento del numero di capezzoli e ghiandole mammarie, rispettivamente.
FILOGENESI SCHELETRICA
I cordati inferiori vivono nell'acqua, mentre i cordati superiori vivono principalmente sulla terra. Il passaggio da un ambiente acquatico più denso a un ambiente aereo ha portato a una significativa complicazione dello scheletro e dei muscoli, in particolare, lo scheletro degli arti e le loro cinture sono diventati più complessi e il numero di muscoli specializzati che servono gli arti è aumentato. Le principali funzioni dello scheletro: supporto, base per l'attaccamento muscolare, partecipazione al metabolismo minerale.
Lo scheletro assile si basa sulla notocorda, quindi le vertebre si sono formate attorno alla notocorda, si sono differenziate, il numero di parti della colonna vertebrale è aumentato, nell'uomo, a causa della postura eretta, si sono formate curve - 2 cifosi e 2 lordosi, e i resti di la notocorda era conservata solo nelle sezioni centrali dei dischi intervertebrali.
Così, nel pesce Ci sono due sezioni della colonna vertebrale:
Tronco (vertebre del tronco articolate con le costole);
Coda;
negli anfibi :
Vengono aggiunte le sezioni cervicale e sacrale (1 vertebra ciascuna);
nei rettili :
Cervicale (8 vertebre, I - atlante, II - asse);
Toracico (articolato con le costole, alcune delle quali articolate con lo sterno davanti);
Lombare, sacrale (contengono un numero maggiore di vertebre);
Coda;
negli uccelli :
Cervicale (fino a 25 vertebre);
Petto;
L'osso sacro lombare e parte delle vertebre caudali sono saldamente collegati, formando un potente sacro;
nei mammiferi :
Cervicale (7 vertebre);
Toracico (9-24 vertebre);
Lombare (3-9 vertebre);
Sacrale (le vertebre si fondono insieme);
Caudale (coccigeo).
Nell'uomo esistono diversi difetti nella formazione del canale spinale, attraverso il quale può emergere la spina bifida, così come la formazione di curve patologiche, comprese quelle laterali (scoliosi), e la regione caudale può essere preservata.
Lo scheletro della testa è diviso in cranio cerebrale e viscerale. Nel processo di evoluzione, si è verificata una diminuzione del numero di ossa del cranio cerebrale, la sostituzione delle parti cartilaginee con ossa e una diminuzione del numero di elementi mobili. Anche il numero degli archi branchiali dello scheletro viscerale è progressivamente diminuito. Il primo e il secondo arco branchiale sono cambiati e hanno dato origine alle mascelle, alle ossa del cranio facciale, alle ossa uditive e alla cartilagine della laringe.
Nei pesci, l'arco I (mascellare, costituito dal palatoquadrato e dalle cartilagini di Meckel) è direttamente collegato all'arco II (costituito da cartilagine iomandibolare e ioide). Questo tipo di connessione tra il cervello e il cranio facciale si chiama iostilo(poiché il II arco si collega alla cartilagine iomandibolare). A partire dagli anfibi il primo arco, con la sua cartilagine palatoquadrata, si fonde con la base del cranio. Questo tipo di connessione si chiama stile automobilistico. La colonna ossea uditiva è formata dalla cartilagine iomandibolare, dalla quale successivamente si forma la staffa. Con disturbi ontogenetici, una persona può avere solo una di queste ossa uditive (come negli anfibi e nei rettili) invece di tre.
Consideriamo la struttura dell'organo uditivo in varie classi di vertebrati. U pescare Esiste solo l'orecchio interno, che comprende il labirinto membranoso con otoliti, canali semicircolari e il rudimento cocleare.
U anfibi C'è un orecchio interno della stessa struttura e un orecchio medio con un osso uditivo e un timpano.
U rettili l'orecchio interno è costituito dalla coclea e dai canali semicircolari, l'orecchio medio è lo stesso degli anfibi, c'è un rudimento del canale uditivo esterno.
U uccelli il canale uditivo esterno è ben formato.
U mammiferi l'orecchio interno comprende la coclea 2,5 giri, canali semicircolari, sacculo, utricolo; orecchio medio - la cavità timpanica in cui si trovano le ossa uditive - la staffa (un residuo della cartilagine iomandibolare), l'incudine (un residuo della cartilagine palato quadrata), il martello (un residuo della cartilagine di Meckel), c'è un timpano membrana, quella esterna comprende il canale uditivo esterno e il padiglione auricolare.
Pertanto, nello scheletro viscerale, nel processo di evoluzione, le mascelle cartilaginee furono sostituite da quelle ossee, cambiò il tipo di attacco delle mascelle e cambiò la funzione degli elementi degli archi viscerali.
Nello scheletro degli arti si formarono cinture con uno scheletro di arti liberi fissato in modo mobile; si formò un arto a cinque dita di tipo terrestre; il numero delle ossa nelle sezioni distali è diminuito; le parti prossimali degli arti furono allungate e le parti distali accorciate.
Una persona può avere i seguenti difetti: polidattilia E polifalange(aumento del numero delle dita e delle falangi, rispettivamente); spostando la cintura degli arti superiori al livello delle I-II vertebre toraciche - La malattia di Sprengel; anomalie congenite delle costole e deformità del torace - acqua ad imbuto, carenato e così via.
FILOGENESI DEI MUSCOLI
I muscoli svolgono la funzione di movimento e si dividono in:
Somatico, che si sviluppa dai miotomi ed è innervato dai nervi spinali (striati);
Viscerale: si sviluppa da altre parti del mesoderma, innervato da nervi autonomi, può essere striato e liscio.
Nei cordati inferiori, i muscoli viscerali si trovano nella regione degli archi branchiali e gradualmente da esso si formano i muscoli della masticazione, i muscoli della faringe, quindi i muscoli del collo e del cingolo scapolare, nonché i muscoli facciali . I muscoli del collo e del viso hanno un'innervazione complessa, associata alle fasi della filogenesi. I muscoli somatici nei cordati inferiori si trovano sotto forma di miomeri, quindi si formano i muscoli degli arti, la forma, le dimensioni e le funzioni dei muscoli diventano più diverse, la loro innervazione aumenta, specialmente gli arti liberi nei primati. I muscoli del tronco si riducono nel processo di evoluzione.
La filogenesi dei sistemi di organi dei cordati è considerata secondo la direzione progressiva dell'evoluzione di questo tipo di animali dal subphylum cranico alla classe dei mammiferi. L'organizzazione dei sistemi di organi della classe degli Uccelli non è descritta a causa del fatto che gli uccelli si sono evoluti dai rettili molto più tardi dei mammiferi e sono un ramo laterale dell'evoluzione dei cordati.
Rivestimenti esterni
Veli Qualsiasi animale svolge sempre la funzione di percepire le irritazioni esterne e protegge anche il corpo dalle influenze dannose dell'ambiente. L'intensificazione della prima funzione del tegumento porta, nel processo di evoluzione degli animali multicellulari, all'emergere di un sistema nervoso e di organi di senso. L'intensificazione della seconda funzione è accompagnata dalla differenziazione. È anche caratteristico un ampliamento delle funzioni, per cui la pelle, come organo protettivo, partecipa anche allo scambio di gas, alla termoregolazione, all'escrezione e all'alimentazione della prole. Ciò è dovuto alla complicazione della struttura degli strati cutanei, all'aspetto e all'ulteriore trasformazione di numerose appendici e ghiandole.
In tutti i cordati la pelle ha una duplice origine: ecto e mesodermica. Dall’ectoderma si sviluppa l’epidermide, dal mesoderma il derma. I senza cranio sono caratterizzati da un debole grado di differenziazione di entrambi gli strati della pelle. L'epidermide è monostrato, cilindrica, contenente ghiandole mucose unicellulari; il derma è lasso e contiene un piccolo numero di cellule del tessuto connettivo.
Nel sottotipo Vertebrato, l'epidermide diventa multistrato, con le cellule nello strato inferiore che si moltiplicano costantemente e negli strati superiori si differenziano, muoiono ed esfoliano. Le fibre del tessuto connettivo compaiono nel derma, conferendo forza al tegumento. La pelle forma appendici, diverse a seconda dello stile di vita e del livello di organizzazione, nonché ghiandole che svolgono varie funzioni.
Nei pesci, le ghiandole dell'epidermide sono unicellulari. Come la lancetta, secernono muco che facilita il movimento nell'acqua. Il corpo del pesce è ricoperto di squame che presentano strutture diverse a seconda della loro posizione sistematica. Vengono chiamate le squame dei pesci cartilaginei placoide. Ha la forma di una punta ed è costituito da dentina ricoperta esternamente da smalto (Fig. 14.1). La dentina è di origine mesodermica; si forma grazie al funzionamento delle cellule del tessuto connettivo che sporgono dall'esterno sotto forma di papilla. Lo smalto, che è una sostanza non cellulare più dura della dentina, è formato dalla papilla dell'epidermide e ricopre l'esterno delle scaglie placoidi.
L'intera superficie del corpo dei pesci cartilaginei, così come la cavità orale, la cui mucosa proviene dall'ectoderma, è ricoperta da scaglie placoidi. Naturalmente, le funzioni delle squame nella cavità orale sono legate alla cattura e alla ritenzione del cibo, quindi sono notevolmente ingrandite e sono denti. I pesci ossei hanno un diverso tipo di squame. Sembrano sottili placche ossee rotonde ricoperte da un sottile strato di epidermide. La scaglia ossea si sviluppa interamente a spese del derma, ma è imparentata in origine con la primitiva scaglia placoide.
La pelle degli anfibi primitivi estinti - gli stegocefali - corrispondeva alla pelle dei pesci ed era anche ricoperta di squame. Gli anfibi moderni hanno una pelle sottile e liscia senza squame, che prende parte allo scambio di gas. Ciò è facilitato dalla presenza di un gran numero di multicellulari ghiandole mucose, il cui segreto idrata costantemente la pelle e ha proprietà battericide. Alcune ghiandole cutanee di numerosi anfibi si sono differenziate in organi produttori di tossine che li proteggono dai nemici (vedere sezione 23.1).
Riso. 14.1. Posa di scaglie placoidi:
1 - cellule che formano lo smalto, 2- epidermide, 3- smalto, 4- scleroblasti che formano dentina, 5- dentina, 6- papilla dermica
I rettili che sono completamente passati all'esistenza terrestre hanno la pelle secca che non partecipa alla respirazione. Strato superiore dell'epidermide cheratinizza. Le scaglie cornee di alcuni rettili sono sottili ed elastiche, mentre in altri si uniscono formando, come nelle tartarughe, un potente guscio corneo. La maggior parte dei rettili fa la muta man mano che cresce, perdendo periodicamente la copertura corneo. I rettili moderni non hanno ghiandole cutanee.
La pelle dei mammiferi è costruita in modo più complesso a causa della varietà di funzioni che svolgono. Sono caratteristici vari derivati della pelle: peli, artigli, corna, zoccoli, nonché ghiandole sudoripare, sebacee e mammarie. I mammiferi più primitivi - insettivori, roditori e alcuni altri - insieme ai capelli, conservavano anche scaglie cornee sulla coda. I loro peli crescono negli spazi tra le squame, in gruppi di 3-7. Nei mammiferi più avanzati che hanno perso le squame, viene mantenuta la stessa disposizione dei peli (Fig. 14.2), che copre quasi tutto il corpo, ad eccezione di alcune aree, ad esempio le piante dei piedi e i palmi degli esseri umani.
Capelli molti mammiferi sono differenziati in tipici, che servono per la termoregolazione, e grandi, o vibrisse, le cui basi sono associate alle terminazioni nervose sensoriali. Nella maggior parte dei mammiferi le vibrisse sono localizzate nella bocca e nel naso; nei primati sono ridotte per l'aumentata funzione tattile degli arti anteriori; in molti ovipari e marsupiali sono sparse in tutto il corpo. Questo fatto potrebbe indicare che i capelli degli antenati dei mammiferi svolgevano principalmente funzioni tattili e poi, con l'aumentare della quantità di capelli, hanno iniziato a prendere parte alla termoregolazione. Durante l'ontogenesi umana si forma un numero maggiore di germogli piliferi, ma alla fine dell'embriogenesi si verifica una riduzione della maggior parte di essi.
Ghiandole sudoripare i mammiferi sono omologhi alle ghiandole cutanee degli anfibi. La loro secrezione può essere mucosa e contenere proteine e grassi. Alcune ghiandole sudoripare si differenziarono nei primi mammiferi in ghiandole mammarie. Negli animali ovipari (ornitorinco, echidna), le ghiandole mammarie sono simili per struttura e sviluppo alle ghiandole sudoripare. Lungo i bordi del capezzolo in via di sviluppo della ghiandola mammaria si possono trovare successive transizioni dalle tipiche ghiandole sudoripare alle ghiandole mammarie (Fig. 14.3). Il numero di ghiandole mammarie e capezzoli è correlato alla fertilità (da 25 a una coppia), ma nell'embriogenesi di tutti i mammiferi si formano "linee lattiginose" sulla superficie addominale, che si estendono dall'ascella all'inguine. Successivamente, i capezzoli si differenziano lungo queste linee, la maggior parte dei quali subiscono poi una riduzione e scompaiono. Pertanto, nell'embriogenesi umana, inizialmente si formano cinque paia di capezzoli e successivamente ne rimane solo uno (Fig. 14.4).

Riso. 14.4. Embriogenesi della parete addominale anteriore umana. UN - embrione a 5 settimane di età (linee lattiginose visibili); B - differenziazione di cinque paia di capezzoli; IN - embrione a 7 settimane di età

Riso. 14.5. Anomalie ataviche dello sviluppo cutaneo.
UN - ipertricosi; B - polimastia
Ghiandole sebacee si formano nella pelle solo nei mammiferi. La loro secrezione, lubrificando il pelo e la superficie della pelle, conferisce loro non bagnabilità ed elasticità.
L'ontogenesi del tegumento e delle appendici cutanee dei mammiferi e dell'uomo riflette la loro evoluzione in base al tipo di arcallassi. In effetti, né i rudimenti delle scaglie cornee caratteristiche dei rettili, né le forme più antiche di appendici cutanee si riassumono nella loro embriogenesi. Allo stesso tempo, nella fase dell'organogenesi secondaria, si sviluppano immediatamente i rudimenti dei follicoli piliferi. Disturbi nell'ontogenesi precoce della pelle umana possono causare la comparsa di alcune malformazioni ataviche minori: ipertricosi (aumento della crescita dei peli), politelia (aumento del numero di capezzoli), polimastia (aumento del numero di ghiandole mammarie) (Fig. 14.5). Tutti sono associati a una violazione della riduzione del numero in eccesso di queste strutture e riflettono la connessione evolutiva dell'uomo con le forme ancestrali più vicine: i mammiferi. Questo è il motivo per cui è impossibile per gli esseri umani e gli altri mammiferi dare alla luce prole con caratteristiche ataviche della pelle caratteristiche di antenati più lontani. Uno dei segni più noti di prematurità nei neonati è l’aumento della crescita dei peli sulla pelle. Poco dopo la nascita, i peli in eccesso solitamente cadono e i follicoli piliferi diventano più piccoli.
SISTEMA MUSCOLARE
La filogenesi della funzione motoria è alla base della progressiva evoluzione degli animali. Pertanto, il livello della loro organizzazione dipende principalmente dalla natura dell'attività motoria, che è determinata dalle caratteristiche dell'organizzazione sistema muscoloscheletrico, ha subito importanti trasformazioni evolutive nel phylum Chordata a causa di cambiamenti negli habitat e cambiamenti nelle forme di locomozione. Infatti, l'ambiente acquatico negli animali privi di esoscheletro prevede movimenti monotoni dovuti alla flessione dell'intero corpo, mentre la vita terrestre è più favorevole al movimento con l'ausilio degli arti.
Consideriamo separatamente l'evoluzione dello scheletro e del sistema muscolare.
Scheletro
Nei cordati scheletro interno. Secondo la sua struttura e funzioni si divide in scheletro assile, scheletro degli arti e testa.
Scheletro assiale
Nel sottotipo Skullless c'è solo scheletro assiale sotto forma di accordo. È costituito da cellule altamente vacuolate, strettamente adiacenti tra loro e ricoperte all'esterno da comuni membrane elastiche e fibrose. L'elasticità della corda è data dal turgore delle sue cellule e dalla robustezza delle membrane. La notocorda si forma nell'ontogenesi di tutti i cordati e negli animali più altamente organizzati svolge non tanto una funzione di supporto, ma morfogenetica, essendo un organo che svolge l'induzione embrionale.
Per tutta la vita dei vertebrati, la notocorda viene conservata solo nei ciclostomi e in alcuni pesci inferiori. In tutti gli altri animali è ridotto. Nell'uomo, nel periodo postembrionale, i rudimenti della corda sono conservati sotto forma di dischi intervertebrali del nucleo polposo. La conservazione di una quantità eccessiva di materiale notocordale quando la sua riduzione è compromessa comporta il rischio di sviluppare tumori nell'uomo - accordo, derivante sulla sua base.
In tutti i vertebrati la notocorda viene gradualmente sostituita vertebre, sviluppandosi dagli sclerotomi dei somiti e viene sostituito funzionalmente colonna vertebrale. Questo è uno degli esempi più chiari di sostituzione d'organo omotopica (vedi § 13.4). La formazione delle vertebre nella filogenesi inizia con lo sviluppo dei loro archi, che coprono il tubo neurale e diventano siti di attacco muscolare. A partire dai pesci cartilaginei, viene rilevata la cartilagine del guscio della notocorda e la crescita delle basi degli archi vertebrali, a seguito della quale si formano i corpi vertebrali. La fusione degli archi vertebrali superiori sopra il tubo neurale forma i processi spinosi e il canale spinale, che racchiude il tubo neurale (Fig. 14.6).

Riso. 14.6. Sviluppo vertebrale. A-fase iniziale; B- prossima fase:
1 -accordo, 2- conchiglia di accordi, 3- archi vertebrali superiori e inferiori, 4- processo spinoso, 5- zone di ossificazione, rudimento 6-notocorda, 7 - corpo cartilagineo vertebrale
La sostituzione della notocorda con la colonna vertebrale - un organo di sostegno più potente e con struttura segmentale - consente di aumentare le dimensioni complessive del corpo e di attivare la funzione motoria. Ulteriori cambiamenti progressivi nella colonna vertebrale sono associati alla sostituzione dei tessuti - la sostituzione del tessuto cartilagineo con l'osso, che si trova nei pesci ossei, nonché alla sua differenziazione in sezioni.
I pesci hanno solo due parti della colonna vertebrale: tronco E coda. Ciò è dovuto al loro movimento nell'acqua dovuto alla flessione del corpo.
Anche gli anfibi acquisiscono cervicale E sacrale dipartimenti, ciascuno rappresentato da una vertebra. Il primo fornisce una maggiore mobilità della testa e il secondo fornisce supporto agli arti posteriori.
Nei rettili si allunga la colonna cervicale, le cui prime due vertebre sono collegate in modo mobile al cranio e forniscono una maggiore mobilità della testa. Appare lombare una sezione ancora debolmente delimitata da quella toracica, ed il sacro è già costituito da due vertebre.
I mammiferi sono caratterizzati da un numero stabile di vertebre nella regione cervicale, pari a 7. Data la grande importanza nel movimento degli arti posteriori, l'osso sacro è formato da 5-10 vertebre. Le regioni lombare e toracica sono chiaramente delimitate l'una dall'altra.
Nei pesci tutte le vertebre del tronco portano costole che non sono fuse tra loro o con lo sterno. Danno al corpo una forma stabile e forniscono supporto ai muscoli che piegano il corpo sul piano orizzontale. Questa funzione delle costole è preservata in tutti i vertebrati che eseguono movimenti serpentini - negli anfibi e nei rettili dalla coda, quindi le loro costole si trovano anche su tutte le vertebre tranne quelle caudali.
Nei rettili, parte delle costole toraciche si fonde con lo sterno, formando il torace, mentre nei mammiferi il torace comprende 12-13 paia di costole.

Riso. 14.7. Anomalie dello sviluppo dello scheletro assile. UN - costole cervicali vestigiali (mostrate dalle frecce); B - Non fusione dei processi spinosi delle vertebre nelle regioni toracica e lombare. Spina bifida
L'ontogenesi dello scheletro assile umano ricapitola le principali fasi filogenetiche della sua formazione: durante il periodo di neurulazione si forma la notocorda, che viene successivamente sostituita da una spina cartilaginea e poi ossea. Un paio di costole si sviluppano sulle vertebre cervicali, toraciche e lombari, dopo di che le costole cervicali e lombari si riducono e le costole toraciche si fondono davanti l'una con l'altra e con lo sterno, formando la gabbia toracica.
L'interruzione dell'ontogenesi dello scheletro assile nell'uomo può essere espressa in difetti di sviluppo atavici come la mancata fusione dei processi spinosi delle vertebre, con conseguente formazione di spinabifida - difetto del canale spinale. In questo caso, le meningi spesso sporgono attraverso il difetto e a spina bifida(Fig. 14.7).
All'età di 1,5-3 mesi. L'embrione umano ha una colonna vertebrale caudale composta da 8-11 vertebre. La violazione della loro riduzione spiega successivamente la possibilità del verificarsi di un'anomalia così nota dello scheletro assile come persistenza della coda.
La violazione della riduzione delle costole cervicali e lombari è alla base della loro conservazione nell'ontogenesi postnatale.
Scheletro della testa
La continuazione anteriore dello scheletro assile è assiale, O cervello, cranio, serve a proteggere il cervello e gli organi sensoriali. Si sviluppa accanto ad esso viscerale, O cranio facciale, formando un supporto per la parte anteriore del tubo digerente. Entrambe le parti del cranio si sviluppano in modo diverso e da rudimenti diversi. Nelle prime fasi dell'evoluzione e dell'ontogenesi non sono collegati tra loro, ma in seguito sorge questa connessione.

Riso. 14.8. Cranio umano con sutura metodica (indicato dalla freccia)
Nella parte posteriore del cranio assiale durante lo sviluppo si riscontrano tracce di segmentazione, pertanto si ritiene che sia il risultato della fusione dell'anlage delle vertebre anteriori tra loro. La composizione del cranio cerebrale comprende anche l'anlage di capsule cartilaginee di origine mesenchimale che circondano gli organi dell'udito, dell'olfatto e della vista. Inoltre, la parte del cranio cerebrale (che si trova anteriormente alla sella turcica), che non presenta segmentazione, si sviluppa apparentemente come una neoplasia a causa dell'aumento delle dimensioni del prosencefalo.
Filogeneticamente, il cranio cerebrale ha attraversato tre fasi di sviluppo: membranoso, cartilagineo E osso.
Nei ciclostomi è quasi interamente membranoso e non ha una parte anteriore non segmentata.
Il cranio dei pesci cartilaginei è quasi interamente cartilagineo e comprende sia la parte posteriore, prevalentemente segmentata, che quella anteriore.
Nei pesci ossei e in altri vertebrati, il cranio assiale diventa osseo a causa dei processi di ossificazione della cartilagine nella zona della sua base (ossa basali, sfenoidi, etmoidali) e per la comparsa di ossa tegumentarie nella sua parte superiore (ossa parietali , ossa frontali, nasali). Le ossa del cranio assiale subiscono oligomerizzazione nel processo di progressiva evoluzione. Ciò è indicato dall'apparizione di un gran numero di zone di ossificazione e dalla loro successiva fusione insieme durante la formazione di ossa come frontale, temporale, ecc. Nell'uomo sono ampiamente conosciute anomalie del cranio cerebrale come la presenza di ossa interparietali, nonché di due ossa frontali con una sutura metopica tra di loro (Fig. 14.8). Non sono accompagnati da alcun fenomeno patologico e quindi vengono solitamente scoperti per caso dopo la morte.
Il cranio viscerale appare per la prima volta anche nei vertebrati inferiori. È formato da mesenchima di origine ectodermica, che è raggruppato sotto forma di condensazioni a forma di arco negli spazi tra le fessure branchiali della faringe. I primi due archi ricevono uno sviluppo particolarmente forte e danno origine agli archi mascellare e ioide degli animali adulti. Gli archi successivi, in numero di 4-5 paia, svolgono una funzione di sostegno per le branchie e vengono chiamati branchie.
Nei pesci cartilaginei, davanti all'arco mascellare di solito ci sono 1-2 paia di archi premascellari, che sono di natura rudimentale. Ciò indica che gli antenati dei vertebrati avevano un numero di archi viscerali maggiore di 6 o 7 e la loro differenziazione avveniva sullo sfondo dell'oligomerizzazione.
L'arco della mascella è costituito da due cartilagini. Si chiama quello in alto palatoquadrato, he svolge la funzione della mascella superiore primaria. Inferiore, o Meckel, cartilagine - mascella inferiore primaria. Sul lato ventrale della faringe le cartilagini di Meckel sono collegate tra loro in modo tale che l'arco mascellare racchiude la cavità orale in un anello. Il secondo arco viscerale su ciascun lato è costituito da iomandibolare cartilagine fusa alla base del cranio e ioide collegato alla cartilagine di Meckel. Pertanto, nei pesci cartilaginei, entrambe le mascelle primarie sono collegate al cranio assiale attraverso un secondo arco viscerale, in cui la cartilagine iomandibolare funge da sospensione al cranio cerebrale. Questo tipo di connessione tra le mascelle e il cranio viene chiamato assiale iostilo(Fig. 14.9).
Nei pesci ossei inizia la sostituzione delle mascelle primarie con quelle secondarie, costituite da false ossa: la mascella e la premascella in alto e il dente in basso. Le cartilagini palatoquadrata e di Meckel diminuiscono di dimensioni e si spostano posteriormente. La cartilagine iomandibolare continua a funzionare come una sospensione, quindi il cranio rimane iostilo.
Gli anfibi, in connessione con il passaggio all'esistenza terrestre, hanno subito cambiamenti significativi nel cranio viscerale. Gli archi branchiali vengono parzialmente ridotti e parzialmente, modificando le loro funzioni, diventano parte dell'apparato cartilagineo della laringe. L'arco mascellare con il suo elemento superiore - la cartilagine quadrata palatina - si fonde completamente con la base del cranio encefalico, e il cranio diventa così stile automobilistico. La cartilagine iomandibolare, notevolmente ridotta e liberata dalla funzione di sospensione, situata nella regione della prima fessura branchiale all'interno della capsula uditiva, ha assunto la funzione di ossicolo uditivo - una colonna - che trasmette le vibrazioni sonore dall'orecchio esterno a quello interno .
Anche il cranio viscerale dei rettili è autostilo. L'apparato mandibolare è caratterizzato da un grado di ossificazione maggiore rispetto a quello degli anfibi. Parte del materiale cartilagineo degli archi branchiali fa parte non solo della laringe, ma anche della trachea.
La mascella inferiore dei mammiferi si articola con l'osso temporale con una complessa articolazione che consente non solo la cattura del cibo, ma anche complessi movimenti di masticazione.
Un ossicolo uditivo - colonna,- caratteristico degli anfibi e dei rettili, diminuendo di dimensioni, si trasforma in staffa, ed i rudimenti delle cartilagini palatoquadrata e di Meckel, uscendo completamente dall'apparato mandibolare, si trasformano, rispettivamente, in incudine E martello. Si crea così un'unica catena funzionale di tre ossicini uditivi nell'orecchio medio, caratteristica solo dei mammiferi (Fig. 14.9).

Riso. 14.9. Evoluzione dei primi due archi branchiali viscerali dei vertebrati.
UN- pesci cartilaginei; B- anfibio; IN- rettile; G- mammifero:
1 -cartilagine palatoquadrata, cartilagine di 2-Meckel, 3- cartilagine iomandibolare, 4-ioide, 5- colonna, 6- ossa sovrapposte delle mascelle secondarie, 7-incudine, 8- staffa, 9- martello; le formazioni omologhe sono indicate dalla corrispondente ombreggiatura
La ricapitolazione delle principali fasi della filogenesi del cranio viscerale avviene anche nell'ontogenesi umana. La violazione della differenziazione degli elementi dell'arco branchiale mascellare negli ossicini uditivi è un meccanismo per la formazione di una tale malformazione dell'orecchio medio come la posizione nella cavità timpanica di un solo ossicolo uditivo - la colonna, che corrisponde all'osso struttura dell'apparato di trasmissione del suono di anfibi e rettili.
Scheletro degli arti
I cordati hanno arti spaiati e accoppiati. Spaiati (pinne dorsali, caudali e anali) sono i principali organi di locomozione negli anfibi, nei pesci e, in misura minore, negli anfibi caudati. I pesci sviluppano anche arti accoppiati - pinne pettorali e pelviche, sulla base dei quali arti accoppiati di terrestri successivamente si sviluppano i quadrupedi.
Diamo uno sguardo più da vicino all'origine e all'evoluzione degli arti accoppiati.
Nelle larve di pesce, così come nei moderni pesci senza cranio, vengono chiamate pieghe laterali della pelle metapleurico(Fig. 14.10). Non hanno né uno scheletro né muscoli propri, svolgono un ruolo passivo: stabilizzano la posizione del corpo e aumentano l'area della superficie addominale, facilitando il movimento nell'ambiente acquatico. Probabilmente, negli antenati dei pesci che passavano ad uno stile di vita più attivo, in queste pieghe apparivano elementi muscolari e raggi cartilaginei, associati in origine ai somiti e quindi localizzati metamericamente. Tali pieghe, avendo acquisito mobilità, possono fungere da timoni di profondità, tuttavia, per cambiare la posizione del corpo nello spazio, le loro sezioni anteriore e posteriore sono di maggiore importanza, poiché sono le più distanti dal centro di gravità. Pertanto, l’evoluzione ha seguito la strada dell’intensificazione delle funzioni delle parti esterne e dell’indebolimento delle funzioni delle parti centrali.

Riso. 14.10. Formazione degli arti anteriori e posteriori dalle pieghe metapleuriche: IO-III-ipotetici stadi dell'evoluzione
Di conseguenza, le pinne pettorali si sono sviluppate dalle sezioni anteriori delle pieghe e le pinne ventrali da quelle posteriori (Fig. 14.10). È possibile che la formazione di sole due paia di arti sui lati laterali del corpo sia stata preceduta dalla disintegrazione di pieghe continue in un numero di pinne pari, di cui anche quella anteriore e quella posteriore erano più importanti. Ciò è evidenziato dall'esistenza di resti fossili dei più antichi pesci poco organizzati e dotati di numerose pinne (Fig. 14.11). A causa della fusione delle basi dei raggi cartilaginei, brachiale E cintura pelvica. Riposo le loro aree si sono differenziate in scheletro degli arti liberi.

Riso. 14.11. Un antico pesce simile a uno squalo con numerosi arti accoppiati
Nella maggior parte dei pesci, lo scheletro delle pinne pari è diviso in una sezione prossimale, costituita da un piccolo numero di placche cartilaginee o ossee, e una sezione distale, che comprende un gran numero di raggi segmentati radialmente. Le pinne sono collegate in modo inattivo alle cinture degli arti. Non possono servire come supporto per il corpo quando si muove sul fondo o sulla terra. Nei pesci con pinne lobate, lo scheletro degli arti accoppiati ha una struttura diversa. Il numero totale dei loro elementi ossei è ridotto e sono di dimensioni maggiori. La sezione prossimale è costituita da un solo elemento osseo di grandi dimensioni, corrispondente all'omero o al femore degli arti anteriori o posteriori. Seguono due ossa più piccole, omologhe all'ulna e al radio o alla tibia e al perone. Su di essi poggiano 7-12 raggi posizionati radialmente. In connessione con le cinture degli arti di tale pinna, sono coinvolti solo gli omologhi dell'omero o del femore, quindi le pinne dei pesci con pinne lobate sono attivamente mobili (Fig. 14.12, A, B) e può essere utilizzato non solo per cambiare la direzione del movimento nell'acqua, ma anche per spostarsi lungo un substrato solido.
La vita di questi pesci in bacini poco profondi in prosciugamento nel periodo devoniano ha contribuito alla selezione di forme con arti più sviluppati e mobili. La presenza in essi di ulteriori organi respiratori (vedi Sezione 14.3.4) divenne il secondo prerequisito per raggiungere la terra e l'emergere di altri adattamenti all'esistenza terrestre, che portarono all'origine degli anfibi e dell'intero gruppo Tetrapoda. I loro primi rappresentanti - gli stegocefali - avevano arti a sette e cinque dita che conservavano somiglianze con le pinne dei pesci con pinne lobate (Fig. 14.12, B)

Riso. 14.12. Scheletro dell'arto di un pesce con pinne lobate ( UN), la sua base ( B) e lo scheletro della zampa anteriore di Stegocefalo ( IN):IO- omero, 2-ulna, 3- raggio
Nello scheletro del polso viene preservata la corretta disposizione radiale degli elementi ossei in 3-4 file, nel metacarpo ci sono 7-5 ossa, e quindi anche le falangi di 7-5 dita giacciono radialmente.
Negli anfibi moderni, il numero di dita negli arti è cinque oppure sono oligomerizzate a quattro.
Un'ulteriore trasformazione progressiva degli arti si esprime in un aumento del grado di mobilità delle articolazioni ossee, in una diminuzione del numero di ossa nel polso, prima a tre file negli anfibi e poi a due nei rettili e nei mammiferi. Allo stesso tempo diminuisce anche il numero delle falangi delle dita. Sono caratteristici anche l'allungamento delle parti prossimali dell'arto e l'accorciamento delle parti distali.
Anche la disposizione degli arti cambia durante l'evoluzione. Se nei pesci le pinne pettorali sono a livello della prima vertebra e sono rivolte ai lati, allora nei vertebrati terrestri, a causa della complicazione dell'orientamento nello spazio, appare un collo e si verifica la mobilità della testa, e nei rettili e soprattutto Nei mammiferi, a causa dell'elevazione del corpo dal suolo, gli arti anteriori si spostano posteriormente e sono orientati non orizzontalmente, ma verticalmente. Lo stesso vale per gli arti posteriori.
La varietà delle condizioni di vita fornite dallo stile di vita terrestre offre una varietà di forme di movimento: saltare, correre, strisciare, volare, scavare, arrampicarsi su rocce e alberi e, quando si ritorna nell'ambiente acquatico, nuotare. Pertanto, nei vertebrati terrestri si può trovare sia una varietà quasi illimitata di arti sia la loro completa riduzione secondaria, e molti adattamenti simili degli arti in vari ambienti si sono ripetutamente verificati in modo convergente (Fig. 14.13). Tuttavia, durante il processo di ontogenesi, la maggior parte dei vertebrati terrestri presenta caratteristiche comuni nello sviluppo degli arti: la formazione dei primordi sotto forma di pieghe scarsamente differenziate, la formazione di primordi a sei o sette dita nelle mani e nei piedi, le parti più esterne che presto si riducono e successivamente si sviluppano solo cinque (Fig. 14.14 ).

Riso. 14.13. Scheletro della zampa anteriore dei vertebrati terrestri. UN-rana- B-salamandra; IN-coccodrillo; G-pipistrello; D-Umano: 1 -omero, osso di 2 raggi, 3 - ossa del polso, 4 - metacarpo, 5 - falangi delle dita, 6 -osso del gomito
Riso. 14.14. Struttura di un arto vertebrato in via di sviluppo: pp - prepollex, pin - postminimus - dita rudimentali aggiuntive I e VII
È interessante notare che nell'embriogenesi dei vertebrati superiori si ricapitola non solo la struttura degli arti degli antenati, ma anche il processo della loro eterotopia. Pertanto, negli esseri umani, gli arti superiori si formano a livello delle vertebre cervicali 3a-4a e quelli inferiori a livello delle vertebre lombari. Allo stesso tempo, gli arti ricevono innervazione dalle parti corrispondenti del midollo spinale. L'eterotopia degli arti è accompagnata dalla formazione dei plessi nervosi cervicale, lombare e sacrale, i cui nervi sono collegati, da un lato, a quei segmenti del midollo spinale da cui sono cresciuti al momento della formazione del arti e, dall'altro, agli arti che si sono spostati in una nuova posizione (Fig. 14.15; vedere anche sezione 14.2.2.2).
Nell'ontogenesi umana sono possibili numerosi disturbi che portano alla formazione di malformazioni congenite ataviche degli arti. COSÌ, polidattilia, oppure un aumento del numero delle dita, ereditato come carattere autosomico dominante, è il risultato dello sviluppo degli anlage di dita aggiuntive, che normalmente sono caratteristici di forme ancestrali lontane. È noto il fenomeno della polifalange, caratterizzato da un aumento del numero delle falangi, solitamente il pollice. La sua presenza si basa sullo sviluppo di tre falangi nel primo dito, come normalmente si osserva nei rettili e negli anfibi con dita indifferenziate. La polifalange bilaterale viene ereditata con modalità autosomica dominante.
Una malformazione grave è una violazione dell'eterotopia del cingolo degli arti superiori dalla regione cervicale al livello della 1a-2a vertebra toracica. Questa anomalia si chiama La malattia di Sprengel o posizione alta congenita della scapola (Fig. 14.16). Si esprime nel fatto che il cingolo scapolare su uno o entrambi i lati è più alto di diversi centimetri rispetto alla posizione normale. A causa del fatto che un tale disturbo è spesso accompagnato da anomalie delle costole, della colonna vertebrale toracica e dalla deformazione delle scapole, si dovrebbe pensare che i meccanismi della sua insorgenza non siano solo una violazione del movimento degli organi, ma anche una conseguente violazione delle correlazioni morfogenetiche (vedi § 13.4).
Una revisione anatomica comparativa dell'evoluzione dello scheletro cordato indica che lo scheletro umano è completamente omologo all'apparato di sostegno delle forme ancestrali e affini. Pertanto, molte malformazioni del suo sviluppo nell'uomo possono essere spiegate dalla relazione dei mammiferi con rettili, anfibi e pesci. Tuttavia, nel processo di antropogenesi, sono apparse caratteristiche scheletriche che sono caratteristiche solo degli esseri umani e sono associate alla loro postura eretta e all'attività lavorativa. Questi includono: 1) cambiamenti nel piede che ha cessato di svolgere una funzione di presa, espresso nella perdita della capacità di opporsi all'alluce e nell'aspetto dei suoi archi, che servono per assorbire gli urti quando si cammina; 2) cambiamenti nella colonna vertebrale - la sua curva a forma di S, che fornisce plasticità ai movimenti in posizione verticale; 3) cambiamenti nel cranio: una forte diminuzione della sua parte facciale e un aumento del cervello, spostamento anteriore del forame magno, un aumento del processo mastoideo e levigatura del rilievo occipitale, a cui sono collegati i muscoli del collo e il legamento nucale allegato; 4) specializzazione degli arti superiori (anteriori) come organo del lavoro; 5) la comparsa di una sporgenza del mento in connessione con lo sviluppo del linguaggio articolato.

Riso. 14.15. Formazione degli arti anteriori, loro eterotopia e innervazione nell'ontogenesi umana. UN-crescita dei miotomi cervicali nella zampa anteriore in via di sviluppo dell'embrione; B-sviluppo dell'innervazione cutanea della mano; IN- localizzazione dei plessi cervicale e brachiale coinvolti nell'innervazione del braccio:
1 -miotomi cervicali, 2- miotomi toracici, 3 -miotomi lombari; le lettere C, T, L indicano i segmenti cervicale, toracico e lombare

Riso. 14.16. Malattia di Sprengel (vedi testo per la spiegazione)
Nonostante la formazione delle caratteristiche anatomiche e morfologiche dello scheletro umano sia apparentemente completata, gli adattamenti alla deambulazione eretta nell'uomo sono, come tutti gli adattamenti in generale, di natura relativa. Pertanto, con un'attività fisica intensa, è possibile lo spostamento delle vertebre o dei dischi intervertebrali. Passando alla camminata eretta, l'uomo ha perso la capacità di correre velocemente e si muove molto più lentamente della maggior parte degli animali a quattro zampe.
Naturalmente, durante lo sviluppo intrauterino, le caratteristiche scheletriche che caratterizzano una persona come specie biologica unica si formano nelle sue fasi finali o addirittura, come la colonna vertebrale a forma di S, nel primo periodo di sviluppo postnatale. In realtà sono anabolizzanti sorti durante la filogenesi dei primati. Ecco perché anomalie ataviche lo scheletro associato a ritardi nello sviluppo di caratteristiche caratteristiche solo degli esseri umani sono i più comuni. Praticamente non riducono la vitalità, ma i bambini con loro necessitano di correzione ortopedica, ginnastica e massaggio. Tali anomalie includono forme lievi di piede piatto congenito, piede torto, torace stretto, assenza di sporgenza del mento e alcune altre.
Sistema muscolare
Nei rappresentanti del phylum Chordata, i muscoli sono divisi in base alla natura dello sviluppo e dell'innervazione in somatici e viscerali.
Muscolatura somatica si sviluppa dai miotomi ed è innervato da nervi, le cui fibre escono dal midollo spinale come parte delle radici ventrali dei nervi spinali. Muscoli viscerali si sviluppa da altre parti del mesoderma ed è innervato dai nervi del sistema nervoso autonomo. Tutti i muscoli somatici sono striati e i muscoli viscerali possono essere striati o lisci (Fig. 14.17).

Riso. 14.17. Muscoli somatici e viscerali dei vertebrati:
1 -muscoli somatici che si sviluppano dai miotomi, 2- muscoli viscerali della regione branchiale
Muscoli viscerali
I cambiamenti più significativi hanno subito i muscoli viscerali associati agli archi viscerali della parte anteriore del tubo digerente. Nei vertebrati inferiori, la maggior parte di questo muscolo è rappresentata dal costrittore generale dell'apparato viscerale - m. constrictor superficialis, che copre l'intera area degli archi branchiali su tutti i lati. Nella zona dell'arco mascellare questo muscolo è innervato nervo trigemino(V), nella zona dell'arco ioide - facciale(VII), nella zona del primo arco branchiale - glossofaringeo(IX), infine, la sua parte situata più caudalmente - vagabondaggio nervo (X). A questo proposito, tutti i derivati dei corrispondenti archi viscerali e i muscoli ad essi associati vengono successivamente innervati in tutti i vertebrati dai nervi elencati.
Nella parte anteriore del compressore è presente una grossa massa muscolare che serve l'apparato mandibolare. Dietro l'apparato viscerale è differenziato il muscolo trapezio m. trapezio, attaccato in fasci separati alle ultime fessure branchiali e al bordo anteriore della parte dorsale del cingolo scapolare. Parte del costrittore superficiale nell'area dell'arco ioide nei rettili cresce, copre il collo dal basso e dai lati e forma il costrittore cervicale m. sfintere del collo. Nei mammiferi, questo muscolo è diviso in due strati: profondo e superficiale. Quello profondo mantiene lo stesso nome, mentre quello superficiale si chiama platysma myoides e si trova sottocute. Questi due muscoli crescono su tutta la regione della testa e danno origine ad un complesso sistema di muscoli sottocutanei facciali, che nei primati e nell'uomo viene chiamato imitare. Pertanto, tutti imitano
LEZIONE MIOLOGIA FILOGENESI, ONTOGENESI E ANATOMIA FUNZIONALE DEL SISTEMA MUSCOLARE Eseguita da: Vladimirova Ya. B. Kokoreva T. V.

 Muscoli o muscoli (dal latino musculus - topo, piccolo topo) sono organi del corpo di animali e esseri umani, costituiti da tessuto muscolare elastico, elastico, capace di contrarsi sotto l'influenza degli impulsi nervosi. Progettato per eseguire varie azioni: movimenti del corpo, contrazione delle corde vocali, respirazione. I muscoli sono costituiti per l’86,3% da acqua. Nel corpo umano ci sono 640 muscoli
Muscoli o muscoli (dal latino musculus - topo, piccolo topo) sono organi del corpo di animali e esseri umani, costituiti da tessuto muscolare elastico, elastico, capace di contrarsi sotto l'influenza degli impulsi nervosi. Progettato per eseguire varie azioni: movimenti del corpo, contrazione delle corde vocali, respirazione. I muscoli sono costituiti per l’86,3% da acqua. Nel corpo umano ci sono 640 muscoli
 Motivazione: - - - possibilità del movimento eseguito, volume del movimento; i movimenti attivi o passivi sono attivati dall'uno o dall'altro gruppo muscolare; influenzando il sistema muscolare, modifichiamo la condizione generale; il sollievo muscolare è una guida per la topografia dei vasi sanguigni e dei nervi; trapianto muscolare, cioè il muscolo può essere “reimparato”.
Motivazione: - - - possibilità del movimento eseguito, volume del movimento; i movimenti attivi o passivi sono attivati dall'uno o dall'altro gruppo muscolare; influenzando il sistema muscolare, modifichiamo la condizione generale; il sollievo muscolare è una guida per la topografia dei vasi sanguigni e dei nervi; trapianto muscolare, cioè il muscolo può essere “reimparato”.
 Sviluppo dei muscoli di origine cranica - dai miotomi della testa (sclerotomi) e dal mesenchima degli archi branchiali. Innervati da rami dei nervi cranici di origine spinale - dai miotomi del tronco dell'embrione: dai miotomi ventrali sono innervati dai rami anteriori del SMN; - dai miotomi dorsali sono innervati dai rami posteriori dei SMN - Muscoli Autoctoni - muscoli che rimangono nella sede del loro anlage primario. I muscoli troncofughi sono muscoli che si sono spostati dal tronco agli arti. I muscoli troncopeti sono muscoli che si sono spostati dagli arti al busto.
Sviluppo dei muscoli di origine cranica - dai miotomi della testa (sclerotomi) e dal mesenchima degli archi branchiali. Innervati da rami dei nervi cranici di origine spinale - dai miotomi del tronco dell'embrione: dai miotomi ventrali sono innervati dai rami anteriori del SMN; - dai miotomi dorsali sono innervati dai rami posteriori dei SMN - Muscoli Autoctoni - muscoli che rimangono nella sede del loro anlage primario. I muscoli troncofughi sono muscoli che si sono spostati dal tronco agli arti. I muscoli troncopeti sono muscoli che si sono spostati dagli arti al busto.



 Striato Liscio 1. L'unità organizzativa è il miocita. Lunghezza circa 50 micron. Larghezza da 6 micron. 2. Contrazione involontaria Controllo da parte del sistema nervoso autonomo Il movimento è ondulatorio, funziona lentamente, poiché la fibra nervosa NON si avvicina a ogni cellula Entrano in azione lentamente, ma rimangono a lungo Non hanno un esatto orientamento spaziale delle cellule 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiaco L'unità di organizzazione è la fibra muscolare, un insieme di mioblasti fluttuanti nel citoplasma comune (sarcoplasma). Hanno un sarcolemma comune. Lunghezza circa 40 -100 mm. Larghezza da 7 mm. Contrazione volontaria Controllo da parte del sistema nervoso somatico Contrazione rapida, reazione rapida, quindi ogni fibra muscolare ha una sinapsi neuromuscolare Si attiva rapidamente, ma ha un effetto a breve termine Chiaro orientamento delle fibre muscolari
Striato Liscio 1. L'unità organizzativa è il miocita. Lunghezza circa 50 micron. Larghezza da 6 micron. 2. Contrazione involontaria Controllo da parte del sistema nervoso autonomo Il movimento è ondulatorio, funziona lentamente, poiché la fibra nervosa NON si avvicina a ogni cellula Entrano in azione lentamente, ma rimangono a lungo Non hanno un esatto orientamento spaziale delle cellule 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiaco L'unità di organizzazione è la fibra muscolare, un insieme di mioblasti fluttuanti nel citoplasma comune (sarcoplasma). Hanno un sarcolemma comune. Lunghezza circa 40 -100 mm. Larghezza da 7 mm. Contrazione volontaria Controllo da parte del sistema nervoso somatico Contrazione rapida, reazione rapida, quindi ogni fibra muscolare ha una sinapsi neuromuscolare Si attiva rapidamente, ma ha un effetto a breve termine Chiaro orientamento delle fibre muscolari

 Tra le fibre muscolari ci sono sottili strati di tessuto connettivo fibroso sciolto: l'endomisio. In essa sono intrecciate le fibre di collagene dello strato esterno della membrana basale, il che aiuta a combinare le forze durante la contrazione dei miosimplasti. Strati più spessi di tessuto connettivo lasso circondano diverse fibre muscolari, formando il perimisio e dividendo il muscolo in fasci. Diversi fasci sono riuniti in gruppi più grandi, separati da strati di tessuto connettivo più spessi. Il tessuto connettivo che circonda la superficie del muscolo è chiamato epimisio.
Tra le fibre muscolari ci sono sottili strati di tessuto connettivo fibroso sciolto: l'endomisio. In essa sono intrecciate le fibre di collagene dello strato esterno della membrana basale, il che aiuta a combinare le forze durante la contrazione dei miosimplasti. Strati più spessi di tessuto connettivo lasso circondano diverse fibre muscolari, formando il perimisio e dividendo il muscolo in fasci. Diversi fasci sono riuniti in gruppi più grandi, separati da strati di tessuto connettivo più spessi. Il tessuto connettivo che circonda la superficie del muscolo è chiamato epimisio.
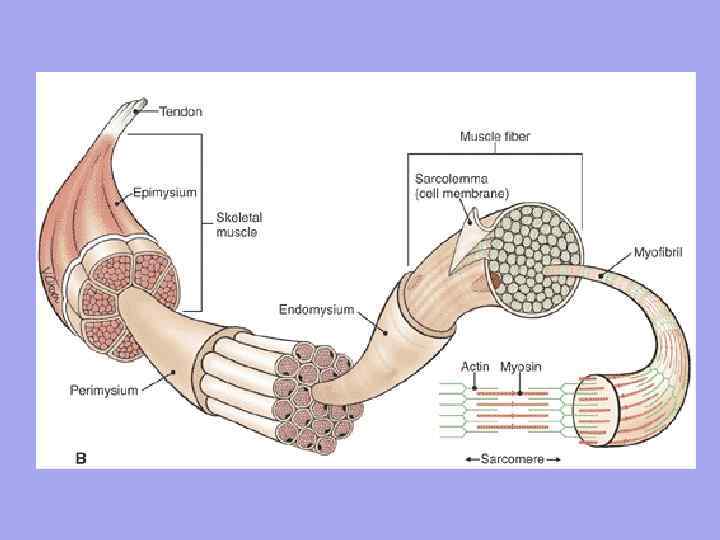
 Il muscolo come organo contiene tessuto connettivo. L'endomisio è un sottile tessuto connettivo che circonda ciascuna fibra muscolare e piccoli gruppi di fibre. Perimisio: copre complessi più grandi di fibre muscolari e fasci muscolari.
Il muscolo come organo contiene tessuto connettivo. L'endomisio è un sottile tessuto connettivo che circonda ciascuna fibra muscolare e piccoli gruppi di fibre. Perimisio: copre complessi più grandi di fibre muscolari e fasci muscolari.
 Significato dell'endomisio e del perimisio 1. Attraverso l'endomisio e il perimisio, vasi e nervi si avvicinano alla fibra muscolare. Formano lo stroma dell'organo; 2. Le fibre muscolari si formano in fasci, i fasci in muscoli; 3. Poiché l'endomisio è fuso con il sarcolemma della fibra muscolare, la fibra muscolare che si contrae può allungarsi solo fino a un certo limite
Significato dell'endomisio e del perimisio 1. Attraverso l'endomisio e il perimisio, vasi e nervi si avvicinano alla fibra muscolare. Formano lo stroma dell'organo; 2. Le fibre muscolari si formano in fasci, i fasci in muscoli; 3. Poiché l'endomisio è fuso con il sarcolemma della fibra muscolare, la fibra muscolare che si contrae può allungarsi solo fino a un certo limite
 Le miofibrille nella fibra sono circondate da un guscio - sarcolemma e immerse in un mezzo speciale - sarcoplasma. A seconda del pigmento e del contenuto di ossigeno, le fibre si dividono in bianche e rosse. Le fibre bianche sono anaerobiche, contengono più miofibrille e meno sarcoplasma. Si avviano rapidamente, ma non possono funzionare a lungo. Esempio: muscoli sternocleidomastoideo, gastrocnemio. Le fibre rosse sono fibre spesse. C'è molta mioglobina nel sarcoplasma e citocromo nei mitocondri, ma meno miofibrille. Lento all'avvio, ma dura a lungo. Esempio: muscoli della schiena, diaframma.
Le miofibrille nella fibra sono circondate da un guscio - sarcolemma e immerse in un mezzo speciale - sarcoplasma. A seconda del pigmento e del contenuto di ossigeno, le fibre si dividono in bianche e rosse. Le fibre bianche sono anaerobiche, contengono più miofibrille e meno sarcoplasma. Si avviano rapidamente, ma non possono funzionare a lungo. Esempio: muscoli sternocleidomastoideo, gastrocnemio. Le fibre rosse sono fibre spesse. C'è molta mioglobina nel sarcoplasma e citocromo nei mitocondri, ma meno miofibrille. Lento all'avvio, ma dura a lungo. Esempio: muscoli della schiena, diaframma.
 Ogni muscolo ha una rete di vasi sanguigni. Le contrazioni muscolari favoriscono il flusso sanguigno. In un muscolo rilassato e non attivo, la maggior parte dei capillari sanguigni sono chiusi al flusso sanguigno. Quando un muscolo si contrae, tutti i capillari sanguigni si aprono immediatamente.
Ogni muscolo ha una rete di vasi sanguigni. Le contrazioni muscolari favoriscono il flusso sanguigno. In un muscolo rilassato e non attivo, la maggior parte dei capillari sanguigni sono chiusi al flusso sanguigno. Quando un muscolo si contrae, tutti i capillari sanguigni si aprono immediatamente.

 La struttura di un muscolo Ogni muscolo è collegato da un'estremità a un osso (l'origine del muscolo) e dall'altra estremità all'altro (l'attaccatura del muscolo). Il muscolo è diviso in: testa, addome e coda.
La struttura di un muscolo Ogni muscolo è collegato da un'estremità a un osso (l'origine del muscolo) e dall'altra estremità all'altro (l'attaccatura del muscolo). Il muscolo è diviso in: testa, addome e coda.
 Le fibre nervose motorie si avvicinano a ciascuna fibra muscolare e le fibre nervose sensoriali si allontanano.Il numero di terminazioni nervose in un muscolo dipende dal grado di attività funzionale dei muscoli.
Le fibre nervose motorie si avvicinano a ciascuna fibra muscolare e le fibre nervose sensoriali si allontanano.Il numero di terminazioni nervose in un muscolo dipende dal grado di attività funzionale dei muscoli.

 Ogni fibra muscolare è innervata in modo indipendente ed è circondata da una rete di emocapillari, che formano un complesso chiamato myon. Un gruppo di fibre muscolari innervate da un motoneurone è chiamato unità motoria. È caratteristico che le fibre muscolari appartenenti a un'unità motoria non si trovano una accanto all'altra, ma si trovano a mosaico tra le fibre appartenenti ad altre unità.
Ogni fibra muscolare è innervata in modo indipendente ed è circondata da una rete di emocapillari, che formano un complesso chiamato myon. Un gruppo di fibre muscolari innervate da un motoneurone è chiamato unità motoria. È caratteristico che le fibre muscolari appartenenti a un'unità motoria non si trovano una accanto all'altra, ma si trovano a mosaico tra le fibre appartenenti ad altre unità.


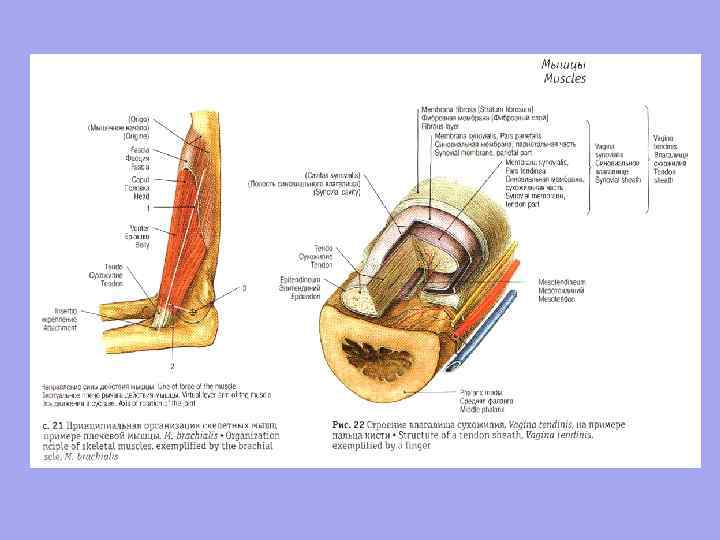
 Un tendine è un cordone di tessuto connettivo fibroso denso che collega un muscolo o si attacca allo scheletro.
Un tendine è un cordone di tessuto connettivo fibroso denso che collega un muscolo o si attacca allo scheletro.
 peritenonio fibre di collagene di tipo IV endotenonio Le fibre di collagene del tendine, intrecciate con le fibre di collagene del periostio, sono intrecciate nella sostanza fondamentale del tessuto osseo, formando creste, tubercoli, tubercoli, depressioni e depressioni sulle ossa.
peritenonio fibre di collagene di tipo IV endotenonio Le fibre di collagene del tendine, intrecciate con le fibre di collagene del periostio, sono intrecciate nella sostanza fondamentale del tessuto osseo, formando creste, tubercoli, tubercoli, depressioni e depressioni sulle ossa.
 La fascia è costituita da fibre di collagene del tessuto connettivo con una piccola aggiunta di fibre elastiche Fascia temporale superficiale Fascia profonda della coscia
La fascia è costituita da fibre di collagene del tessuto connettivo con una piccola aggiunta di fibre elastiche Fascia temporale superficiale Fascia profonda della coscia
 1. 2. 3. 4. 5. La fascia separa i muscoli dalla pelle ed elimina lo spostamento della pelle durante i movimenti di contrazione dei muscoli. La fascia conserva la forza della contrazione muscolare eliminando l'attrito tra i muscoli durante la contrazione. La fascia allunga le grandi vene sotto tensione, a seguito della quale il sangue dalla periferia viene “risucchiato” in queste vene. La fascia è importante come barriera che impedisce la diffusione di infezioni e tumori. Durante le operazioni, la fascia aiuta a determinare la posizione di muscoli, vasi sanguigni e visceri.
1. 2. 3. 4. 5. La fascia separa i muscoli dalla pelle ed elimina lo spostamento della pelle durante i movimenti di contrazione dei muscoli. La fascia conserva la forza della contrazione muscolare eliminando l'attrito tra i muscoli durante la contrazione. La fascia allunga le grandi vene sotto tensione, a seguito della quale il sangue dalla periferia viene “risucchiato” in queste vene. La fascia è importante come barriera che impedisce la diffusione di infezioni e tumori. Durante le operazioni, la fascia aiuta a determinare la posizione di muscoli, vasi sanguigni e visceri.
 Classificazione dei muscoli I muscoli scheletrici variano per forma, struttura, posizione rispetto agli assi delle articolazioni, ecc., e quindi sono classificati in modo diverso.
Classificazione dei muscoli I muscoli scheletrici variano per forma, struttura, posizione rispetto agli assi delle articolazioni, ecc., e quindi sono classificati in modo diverso.

 III. Secondo le caratteristiche funzionali, Statico (forte): ventre corto e tendine lungo. I muscoli lavorano con maggiore forza, ma con un range di movimento minore. Dinamico (abile) – fasci muscolari lunghi, tendini corti. I muscoli lavorano con meno forza, ma producono movimenti più ampi
III. Secondo le caratteristiche funzionali, Statico (forte): ventre corto e tendine lungo. I muscoli lavorano con maggiore forza, ma con un range di movimento minore. Dinamico (abile) – fasci muscolari lunghi, tendini corti. I muscoli lavorano con meno forza, ma producono movimenti più ampi
 Apparato accessorio dei muscoli I muscoli scheletrici sono dotati di un apparato accessorio che ne facilita il funzionamento. n n n Fascia; Guaine osteofasciali; Borse sinoviali; Guaine tendinee sinoviali; Blocchi muscolari; Ossa sesamoidi.
Apparato accessorio dei muscoli I muscoli scheletrici sono dotati di un apparato accessorio che ne facilita il funzionamento. n n n Fascia; Guaine osteofasciali; Borse sinoviali; Guaine tendinee sinoviali; Blocchi muscolari; Ossa sesamoidi.
 Le anomalie dello sviluppo muscolare sono molto comuni e si dividono in tre gruppi: 1. Assenza di qualsiasi muscolo; 2. La presenza di un muscolo aggiuntivo che non esiste in natura. 3. Fasci aggiuntivi di muscoli esistenti.
Le anomalie dello sviluppo muscolare sono molto comuni e si dividono in tre gruppi: 1. Assenza di qualsiasi muscolo; 2. La presenza di un muscolo aggiuntivo che non esiste in natura. 3. Fasci aggiuntivi di muscoli esistenti.
 Difetti dello sviluppo: Sottosviluppo del muscolo sternocleidomastoideo - Torcicollo; Sottosviluppo del diaframma. Causa dell'ernia diaframmatica. Sottosviluppo dei muscoli deltoide e trapezio – Deformazione del cingolo scapolare e della spalla
Difetti dello sviluppo: Sottosviluppo del muscolo sternocleidomastoideo - Torcicollo; Sottosviluppo del diaframma. Causa dell'ernia diaframmatica. Sottosviluppo dei muscoli deltoide e trapezio – Deformazione del cingolo scapolare e della spalla


 I. Forma: fusiforme; A forma di nastro; Piatto largo; Serrato; Lungo; n n n Quadrato; Triangolare; Girare; Deltoide; Soleo, ecc.
I. Forma: fusiforme; A forma di nastro; Piatto largo; Serrato; Lungo; n n n Quadrato; Triangolare; Girare; Deltoide; Soleo, ecc.



 II. Nella direzione delle fibre muscolari Con fibre parallele diritte; Con quelli trasversali; Con circolare; Pennato: A. Unipennato; Bipennato; C. Multipennate. B.
II. Nella direzione delle fibre muscolari Con fibre parallele diritte; Con quelli trasversali; Con circolare; Pennato: A. Unipennato; Bipennato; C. Multipennate. B.

 IV. Per funzione: Adduttori; Deviatori; Piegatura; estensore; Pronatori; n n Supporti per arco plantare; filtrare; I muscoli sono sinergici; I muscoli sono antagonisti.
IV. Per funzione: Adduttori; Deviatori; Piegatura; estensore; Pronatori; n n Supporti per arco plantare; filtrare; I muscoli sono sinergici; I muscoli sono antagonisti.
 V. In relazione al giunto: Giunto singolo; Due giunti; Multi-giunto.
V. In relazione al giunto: Giunto singolo; Due giunti; Multi-giunto.
Il sistema muscolare si sviluppa parallelamente allo sviluppo del sistema nervoso come organo esecutivo. Nella filogenesi appare sulla base delle proprietà contrattili degli elementi cellulari degli organismi multicellulari più semplici che rispondono all'irritazione. Inizialmente, nel corpo dell’animale appare il tessuto muscolare liscio, che conserva ancora bene la sua struttura cellulare ed è collegato al sistema nervoso attraverso due neuroni. (La comunicazione efficace con i centri del sistema nervoso viene effettuata attraverso un neurone periferico.) Con il miglioramento delle forme di movimento traslazionale e degli elementi di supporto associati del corpo - appare lo scheletro, appare il tessuto muscolare scheletrico (somatico), che già ha una connessione efferente diretta con il sistema nervoso centrale.
Se negli animali celenterati gli elementi muscolari non sono ancora isolati, ma sono rappresentati da speciali processi muscolari delle cellule epiteliali, che si differenziano in localizzati longitudinalmente (nell'ectoderma) e ad anello (nell'endoderma), quindi nei vermi sono isolati, non sono più associati all'epitelio e formano una borsa muscolocutanea.
Nei cordati, animali più mobili, tutti i muscoli sono divisi in somatici e viscerali. Negli adulti, i muscoli somatici sono striati. Sono rappresentati da uno strato longitudinale di muscoli laterali accoppiati, divisi da membrane in metameri separati (segmenti). La lancetta ha ancora tessuto muscolare liscio nei suoi vasi e nell'intestino. Nei vertebrati, come nella lancetta, i muscoli sono divisi in somatici e lisci, differendo non solo per la struttura, ma anche per la natura delle connessioni nervose.
Quanto più complesso diventa l'organismo dei vertebrati nella filogenesi, tanto più i muscoli scheletrici perdono il loro metamerismo, tanto più si differenziano, dividendosi prima in strati dorsale e ventrale, e poi in strati, sezioni, gruppi muscolari e muscoli separati.
Nei vertebrati terrestri tutti i muscoli scheletrici sono già divisi in gruppi: muscoli del tronco, della testa e degli arti. Il tessuto muscolare liscio rimane ancora principalmente nelle pareti dei vasi sanguigni e degli organi interni. Non importa quanto sia complesso il muscolo scheletrico come organo, è collegato a determinati segmenti del cervello.
Questa connessione viene stabilita fin dai primi momenti della comparsa degli elementi muscolari e non viene interrotta durante le trasformazioni filogenetiche. A questo proposito, è stato stabilito che un gruppo di fibre muscolari che ricevono terminazioni nervose (placche nervose) da una cellula nervosa è un “muscolo unità" . Da quale segmento si sviluppa il muscolo nell'embrione, dallo stesso segmento verrà innervato in futuro. Se un muscolo si muove durante lo sviluppo, segue un nervo (un ottimo esempio di ciò è il diaframma situato dietro la corda toracica). Se il muscolo si differenzia in parti separate, il nervo subisce la stessa differenziazione; se il muscolo si fonde, anche i nervi si fondono. La riduzione del muscolo porta alla riduzione della connessione nervosa. Una connessione così invariabile consente di identificare i percorsi di sviluppo muscolare nell'embriogenesi e nella filogenesi e consente di stabilire attraverso quali segmenti dell'embrione si è formato.
Figura 91. Schema dei muscoli esterni dei vertebrati
Durante il processo di sviluppo individuale (nell'ontogenesi), i muscoli si sviluppano da diverse fonti del mesoderma; 1) a causa del materiale dei miotomi del mesoderma; 2) dal mesoderma non segmentato nella regione branchiale. Pertanto, i muscoli che si sviluppano a causa del mesenchima dell'apparato branchiale sono chiamati viscerali. Inizialmente, nell'embrione, l'anlage degli elementi muscolari, come nella filogenesi, è segmentato e differenziato in metameri separati, separati da partizioni di tessuto connettivo.
Nell'ontogenesi le fasi principali si ripetono, soprattutto nel primo periodo della filogenesi (Fig. 91). Inizialmente, il muscolo laterale è disposto lungo il corpo come una corda longitudinale accoppiata, segmentata da setti di tessuto connettivo in miotomi posizionati metamericamente e ciascun miotomo riceve il nervo di un determinato segmento nervoso. Quindi appare un setto longitudinale di tessuto connettivo, che divide il miotomo in parti dorsale e ventrale. Successivamente, al posto di questo setto longitudinale, si sviluppano i processi trasversali delle vertebre e delle costole. Nella stessa sequenza avviene la divisione dei rami nervosi, mantenendo le connessioni con gli stessi segmenti nervosi. A causa del materiale delle parti dorsali dei miotomi, si sviluppano successivamente i muscoli della colonna vertebrale e, a causa delle parti ventrali, si sviluppano i muscoli delle pareti toraciche laterali e addominali.
Un po' più tardi, i fasci muscolari crescono negli abbozzi degli arti dalle sezioni dorsale e ventrale dei miotomi (da cinque miotomi negli arti pettorali e da otto negli arti pelvici). Con la differenziazione degli strati muscolari in singoli muscoli, i muscoli scheletrici diventano ancora più complessi, soprattutto con la separazione dei muscoli degli arti, anch'essi innervati principalmente dai rami ventrali dei nervi spinali.
I muscoli della testa e alcuni muscoli del collo si sviluppano dal mesoderma non segmentato della regione dei primi cinque archi branchiali. Dai rudimenti del primo arco branchiale si sviluppano i muscoli masticatori associati alla V coppia di nervi cranici, dal secondo i muscoli facciali associati alla VII coppia di nervi cranici.
Il tessuto muscolare, grazie alla sua capacità di contrarsi durante il processo di filogenesi, si è formato in organi separati - muscoli - motori a forma di cintura con forza in grado di sollevare pesi pesanti, sostenere il peso corporeo sulle gambe, sferrare colpi, sviluppare alta velocità durante il movimento , eccetera.
I muscoli, quando si contraggono, non solo provocano il movimento (eseguono un lavoro dinamico), ma forniscono anche il tono muscolare, rafforzando le articolazioni con un certo angolo di combinazione con un corpo fermo, mantenendo una certa postura (eseguono un lavoro statico). Il lavoro statico, soprattutto negli ungulati di grandi dimensioni (pesanti), è estremamente difficile.
Un certo tono muscolare svolge anche una funzione antigravitazionale. Insieme ai tendini, alla fascia e ai legamenti dello scheletro, i muscoli hanno proprietà elastiche, grazie alle quali sono affidabili ammortizzatori, conferendo agli arti maggiore elasticità, e funzionano anche come micropompe che favoriscono la rimozione di sangue e linfa dagli organi (N. I. Ariinchin, 1987).
In ogni specie di animale, nel processo di evoluzione, i muscoli, sviluppandosi, determinano i loro limiti di forza, velocità di movimento, resistenza e massa, che dipende direttamente dalla specie, dall'età, dal sesso, dalla razza dell'animale e la loro formazione. La massa muscolare rappresenta circa il 42-47% del peso corporeo totale nei bovini e nei cavalli, il 34% nelle pecore e il 31% nei suini. Solo il lavoro (allenamento) dei muscoli aiuta ad aumentarne la massa sia aumentando il diametro delle fibre muscolari (ipertrofia) sia aumentandone il numero (iperplasia).
Il lavoro dei muscoli è strettamente connesso con l'organo dell'equilibrio e, in larga misura, con altri organi di senso. Grazie a questa connessione, i muscoli forniscono equilibrio al corpo, precisione dei movimenti e forza. I muscoli sono un potente generatore della trasformazione dell'energia chimica del cibo in energia termica, dell'energia delle deformazioni elastiche in energia meccanica, che viene utilizzata per muovere il sangue attraverso i vasi ed eccitare gli impulsi afferenti inviati al cervello, nonché la transizione in energia termica.
Lezione di biologia sul tema "Filogenesi del sistema muscolo-scheletrico e nervoso" Filogenesi del sistema muscolare: modelli di sviluppo
Biscotti fatti in casa con panna
Re Bhumibol Adulyadej della Thailandia: re divino con macchina fotografica
Salvadanaio - Domande sulla Bibbia
Ricetta: Frittelle di zucchine - Con formaggio - molto tenere e gustose