Tasksstudying this section:
- This section helps to understand the ways of evolution of the organic world, gradual complication not only structural, but also physiological and biochemical mechanisms, which led to the emergence of modern cranial chordates, including primates and humans;
- in the process of comparison, it is possible to clearly verify the physiological conditionality of the appearance or complication of certain structures, as well as their disappearance, i.e., it is possible to better understand the structure of the human body;
- helps to understand human embryology, because according to the basic biogenetic law formulated in 1866 by Haeckel and Müller: ontogeny is a short and rapid repetition of phylogeny.
Therefore, in embryogenesis, humans are characterized by the appearance of structures, functions, and biochemical reactions that were present in their chordate ancestors. Such repetitions are called recapitulations. In human ontogenesis, the structures of adult fish, amphibians and reptiles usually do not appear, but only their larvae. Moreover, the early stages of development recapitulate more completely than the later ones.
But evolutionary development is not a simple repetition of the structural features of ancestors, but presupposes changes during the formation of organs and systems . This:
- cenogenesis - adaptations of the embryo to a different environment, which are lost in adult forms, for example, gills in a tadpole; provisional organs of amniotes;
- phylembryogenesis - developmental changes that have adaptive significance:
A) anabolia (extension) - adding additional stages, for example, the muscular diaphragm of mammals, feathers of birds;
b) deviation - deviation from the program at the middle stages of morphogenesis, for example, the formation of auditory ossicles, the appearance of an alveolar structure of the lungs instead of a cellular one; the biogenetic law is partially fulfilled in these cases;
V) archallaxis - deviation of ontogenesis at the very beginning, when the biogenetic law is not fulfilled, for example, hair formation begins with a thickening of the epidermis, which is immersed in the corium;
G) heterochrony - change in the time of formation of the structure, for example, early formation of the placental amnion; And heterotopia - changing the location of the structure, for example, the formation of the human shoulder girdle at the level of the III-IV cervical vertebrae, which then moves to the level of the I-II thoracic vertebrae.
With disorders of embryogenesis, adults may develop signs that were present in their distant ancestors - atavisms. If they reduce viability, they are called atavistic(or ancestral from anĉetre) vices. Atavisms arise due to:
Underdevelopment of organs (hypoplasia of the diaphragm, cleft palate - “cleft palate”, etc.);
Persistence (preservation) of embryonic structures (patent ductus ductus, lateral neck fistulas, umbilical fistulas, etc.);
Disturbances in the movement of organs in ontogenesis (pelvic location of organs, cryptorchidism, etc.).
Let's consider location of chordates in the animal world and their classification.
The phylum Chordata unites multicellular deuterostomes of different levels of organization. Moreover, absolutely all chordates, at least at one stage of development, have three characteristics that distinguish them from other animals:
- an axial complex of organs, which is based on the internal skeleton - the notochord, above it - the neural tube, below it - the digestive tube;
- the pharynx is penetrated by pharyngeal slits, which in higher chordates are supported by visceral arches;
- tail section
Additionally, one can note the presence of the portal vein venous system (blood from the gastrointestinal tract passes through the liver) and the structure of the integument of the epidermis and dermis, but the integument of echinoderms has a similar structure.
The phylum Chordata includes four subphyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Larval chordates (Urochordata) - ascidians;
- III. Skullless (Acrania) - lancelets;
- IV. Vertebrates (Vertebrata), which include several classes:
Group Anamnia (lower vertebrates)
- Cyclostomes (Cyclostomata) - lampreys, hagfish - do not have jaws;
- superclass Pisces:
A. class Cartilaginous and armored fish (Chondrichthyes) - sharks, rays, chimeras;
b. class Bony fish (Osteichthyes) - other fish;
- Amphibians (Amphibia):
A. Tailed animals - newts, salamanders;
b. Tailless animals - frogs, toads, tree frogs;
V. Legless ones are caecilians;
Group Amniota (higher vertebrates)
- Reptiles (Reptilia):
A. Scaly - lizards, snakes;
b. Turtles;
V. Crocodiles;
g. Beakheads - hatteria;
- Birds (Aves) - a side branch of evolution, have many specific adaptations
A. Ratites - ostriches;
b. Penguins;
V. Keeleds - other birds;
- Mammals (Mammalia):
A. Oviparous - platypus, echidna;
b. Marsupials - kangaroos;
V. Placentals - other animals and humans.
PHYLOGENESIS OF EXTERNAL INTEGRATION
The main functions of the integument are delimitation and protection of a multicellular organism. The integument of vertebrates acquires numerous additional functions. In chordates, the integument consists of:
Epidermis developing from ectoderm;
Dermis developing from mesoderm.
The directions of evolution are as follows:
The single-layer epidermis (lancelet) becomes multilayered (in vertebrates), the layers differentiate and become keratinized on the outside; derivatives of the epidermis appear - horny scales, claws, hair, etc.
The thin, gelatinous connective tissue of the dermis thickens and gains strength due to an increase in the number of fibers.
The epidermis and dermis have numerous derivatives: chromatophores, scales, horny scutes, shells, plates, beak, claws, feathers, hair, horns, hooves. All chordates have glands. In lower animals these are unicellular mucous glands, starting with fish, poisonous glands appear, then sebaceous glands (in birds), in mammals mammary and sweat glands are added.
A person may have the following disorders of integument ontogenesis: albinism(full) and vitiligo(incomplete) skin depigmentation, alopecia(absence) and hypertrichosis(excessive growth) of hair, hyperkeratosis And pachyonychia- thickening of the epidermis and nails, respectively, polythelia And polymastia- an increase in the number of nipples and mammary glands, respectively.
SKELETAL PHYLOGENESIS
Lower chordates live in water, while higher chordates live mainly on land. Moving from a denser water environment to an air environment led to a significant complication of the skeleton and muscles, in particular, the skeleton of the limbs and their girdles became more complex, and the number of specialized muscles serving the limbs increased. The main functions of the skeleton: support, basis for muscle attachment, participation in mineral metabolism.
The axial skeleton is based on the notochord, then vertebrae formed around the notochord, they differentiated, the number of parts of the spine increased, in humans, due to upright posture, bends were formed - 2 kyphosis and 2 lordosis, and the remnants of the notochord were preserved only in the central sections of the intervertebral discs.
Thus, in fish There are two sections of the spine:
Trunk (trunk vertebrae articulated with the ribs);
Tail;
in amphibians :
The cervical and sacral sections are added (1 vertebra each);
in reptiles :
Cervical (8 vertebrae, I - atlas, II - axis);
Thoracic (articulated with the ribs, some of which are articulated with the sternum in front);
Lumbar, sacral (contain a larger number of vertebrae);
Tail;
in birds :
Cervical (up to 25 vertebrae);
Chest;
The lumbar sacrum and part of the caudal vertebrae are firmly connected, forming a powerful sacrum;
in mammals :
Cervical (7 vertebrae);
Thoracic (9-24 vertebrae);
Lumbar (3-9 vertebrae);
Sacral (vertebrae fuse together);
Caudal (coccygeal).
In humans, there are various defects in the formation of the spinal canal, through which spina bifida can emerge, as well as the formation of pathological curves, including lateral ones (scoliosis), and the caudal region can be preserved.
The head skeleton is divided into the cerebral and visceral skull. In the process of evolution, there was a decrease in the number of bones of the brain skull, replacement of cartilaginous parts with bone, and a decrease in the number of moving elements. The number of branchial arches of the visceral skeleton also progressively decreased. The first and second branchial arches have changed and given rise to the jaws, bones of the facial skull, auditory bones, and cartilage of the larynx.
In fish, arch I (maxillary, consists of the palatoquadrate and Meckel's cartilages) is directly connected to arch II (consists of hyomandibular cartilage and hyoid). This type of connection between the brain and facial skull is called hyostyle(since the II arch connects to the hyomandibular cartilage). Beginning with amphibians, the first arch, with its palatoquadrate cartilage, fuses with the base of the cranium. This type of connection is called autostyle. The auditory bone column is formed from the hyomandibular cartilage, from which the stapes is subsequently formed. With ontogeny disorders, a person may have only one of these auditory bones (as in amphibians and reptiles) instead of three.
Let us consider the structure of the hearing organ in various classes of vertebrates. U fish There is only an inner ear, which includes the membranous labyrinth with otoliths, semicircular canals and the cochlear rudiment.
U amphibians There is an inner ear of the same structure and a middle ear with one auditory bone and an eardrum.
U reptiles the inner ear consists of the cochlea and semicircular canals, the middle ear is the same as in amphibians, there is a rudiment of the external auditory canal.
U birds the external auditory canal is well formed.
U mammals the inner ear includes the cochlea 2.5 turns, semicircular canals, saccule, utricle; middle ear - the tympanic cavity in which the auditory bones are located - the stapes (a remnant of the hyomandibular cartilage), the incus (a remnant of the quadratus palatocartilage), the malleus (a remnant of Meckel's cartilage), there is a tympanic membrane, the external includes the external auditory canal and the auricle.
Thus, in the visceral skeleton, in the process of evolution, cartilaginous jaws were replaced by bone ones, the type of attachment of the jaws changed, and the function of the elements of the visceral arches changed.
In the skeleton of the limbs, belts with a movably attached skeleton of free limbs were formed; a five-fingered limb of the terrestrial type was formed; the number of bones in the distal sections decreased; the proximal parts of the limbs were lengthened and the distal parts were shortened.
A person may have the following defects: polydactyly And polyphalanx(increase in the number of fingers and phalanges, respectively); moving the belt of the upper limbs to the level of the I-II thoracic vertebrae - Sprengel's disease; congenital anomalies of the ribs and chest deformities - funnel-water, keeled and etc.
PHYLOGENESIS OF MUSCLES
Muscles perform the function of movement and are divided into:
Somatic, which develops from myotomes and is innervated by spinal nerves (striated);
Visceral - develops from other parts of the mesoderm, innervated by autonomic nerves, can be striated and smooth.
In lower chordates, the visceral muscles are located in the region of the gill arches, and gradually the muscles of mastication, the muscles of the pharynx, then the muscles of the neck and shoulder girdle, as well as facial muscles, are formed from it. The muscles of the neck and face have complex innervation, which is associated with the stages of phylogenesis. Somatic muscles in lower chordates are located in the form of myomeres, then the muscles of the limbs are formed, the shape, size and functions of the muscles become more diverse, their innervation increases, especially the free limbs in primates. The muscles of the trunk are reduced in the process of evolution.
Tasksstudying this section:
- This section helps to understand the ways of evolution of the organic world, gradual complication not only structural, but also physiological and biochemical mechanisms, which led to the emergence of modern cranial chordates, including primates and humans;
- in the process of comparison, it is possible to clearly verify the physiological conditionality of the appearance or complication of certain structures, as well as their disappearance, i.e., it is possible to better understand the structure of the human body;
- helps to understand human embryology, because according to the basic biogenetic law formulated in 1866 by Haeckel and Müller: ontogeny is a short and rapid repetition of phylogeny.
Therefore, in embryogenesis, humans are characterized by the appearance of structures, functions, and biochemical reactions that were present in their chordate ancestors. Such repetitions are called recapitulations. In human ontogenesis, the structures of adult fish, amphibians and reptiles usually do not appear, but only their larvae. Moreover, the early stages of development recapitulate more completely than the later ones.
But evolutionary development is not a simple repetition of the structural features of ancestors, but presupposes changes during the formation of organs and systems . This:
- cenogenesis - adaptations of the embryo to a different environment, which are lost in adult forms, for example, gills in a tadpole; provisional organs of amniotes;
- phylembryogenesis - developmental changes that have adaptive significance:
A) anabolia (extension) - adding additional stages, for example, the muscular diaphragm of mammals, feathers of birds;
b) deviation - deviation from the program at the middle stages of morphogenesis, for example, the formation of auditory ossicles, the appearance of an alveolar structure of the lungs instead of a cellular one; the biogenetic law is partially fulfilled in these cases;
V) archallaxis - deviation of ontogenesis at the very beginning, when the biogenetic law is not fulfilled, for example, hair formation begins with a thickening of the epidermis, which is immersed in the corium;
G) heterochrony - change in the time of formation of the structure, for example, early formation of the placental amnion; And heterotopia - changing the location of the structure, for example, the formation of the human shoulder girdle at the level of the III-IV cervical vertebrae, which then moves to the level of the I-II thoracic vertebrae.
With disorders of embryogenesis, adults may develop signs that were present in their distant ancestors - atavisms. If they reduce viability, they are called atavistic(or ancestral from anĉetre) vices. Atavisms arise due to:
Underdevelopment of organs (hypoplasia of the diaphragm, cleft palate - “cleft palate”, etc.);
Persistence (preservation) of embryonic structures (patent ductus ductus, lateral neck fistulas, umbilical fistulas, etc.);
Disturbances in the movement of organs in ontogenesis (pelvic location of organs, cryptorchidism, etc.).
Let's consider location of chordates in the animal world and their classification.
The phylum Chordata unites multicellular deuterostomes of different levels of organization. Moreover, absolutely all chordates, at least at one stage of development, have three characteristics that distinguish them from other animals:
- an axial complex of organs, which is based on the internal skeleton - the notochord, above it - the neural tube, below it - the digestive tube;
- the pharynx is penetrated by pharyngeal slits, which in higher chordates are supported by visceral arches;
- tail section
Additionally, one can note the presence of the portal vein venous system (blood from the gastrointestinal tract passes through the liver) and the structure of the integument of the epidermis and dermis, but the integument of echinoderms has a similar structure.
The phylum Chordata includes four subphyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Larval chordates (Urochordata) - ascidians;
- III. Skullless (Acrania) - lancelets;
- IV. Vertebrates (Vertebrata), which include several classes:
Group Anamnia (lower vertebrates)
- Cyclostomes (Cyclostomata) - lampreys, hagfish - do not have jaws;
- superclass Pisces:
A. class Cartilaginous and armored fish (Chondrichthyes) - sharks, rays, chimeras;
b. class Bony fish (Osteichthyes) - other fish;
- Amphibians (Amphibia):
A. Tailed animals - newts, salamanders;
b. Tailless animals - frogs, toads, tree frogs;
V. Legless ones are caecilians;
Group Amniota (higher vertebrates)
- Reptiles (Reptilia):
A. Scaly - lizards, snakes;
b. Turtles;
V. Crocodiles;
g. Beakheads - hatteria;
- Birds (Aves) - a side branch of evolution, have many specific adaptations
A. Ratites - ostriches;
b. Penguins;
V. Keeleds - other birds;
- Mammals (Mammalia):
A. Oviparous - platypus, echidna;
b. Marsupials - kangaroos;
V. Placentals - other animals and humans.
PHYLOGENESIS OF EXTERNAL INTEGRATION
The main functions of the integument are delimitation and protection of a multicellular organism. The integument of vertebrates acquires numerous additional functions. In chordates, the integument consists of:
Epidermis developing from ectoderm;
Dermis developing from mesoderm.
The directions of evolution are as follows:
The single-layer epidermis (lancelet) becomes multilayered (in vertebrates), the layers differentiate and become keratinized on the outside; derivatives of the epidermis appear - horny scales, claws, hair, etc.
The thin, gelatinous connective tissue of the dermis thickens and gains strength due to an increase in the number of fibers.
The epidermis and dermis have numerous derivatives: chromatophores, scales, horny scutes, shells, plates, beak, claws, feathers, hair, horns, hooves. All chordates have glands. In lower animals these are unicellular mucous glands, starting with fish, poisonous glands appear, then sebaceous glands (in birds), in mammals mammary and sweat glands are added.
A person may have the following disorders of integument ontogenesis: albinism(full) and vitiligo(incomplete) skin depigmentation, alopecia(absence) and hypertrichosis(excessive growth) of hair, hyperkeratosis And pachyonychia- thickening of the epidermis and nails, respectively, polythelia And polymastia- an increase in the number of nipples and mammary glands, respectively.
SKELETAL PHYLOGENESIS
Lower chordates live in water, while higher chordates live mainly on land. Moving from a denser water environment to an air environment led to a significant complication of the skeleton and muscles, in particular, the skeleton of the limbs and their girdles became more complex, and the number of specialized muscles serving the limbs increased. The main functions of the skeleton: support, basis for muscle attachment, participation in mineral metabolism.
The axial skeleton is based on the notochord, then vertebrae formed around the notochord, they differentiated, the number of parts of the spine increased, in humans, due to upright posture, bends were formed - 2 kyphosis and 2 lordosis, and the remnants of the notochord were preserved only in the central sections of the intervertebral discs.
Thus, in fish There are two sections of the spine:
Trunk (trunk vertebrae articulated with the ribs);
Tail;
in amphibians :
The cervical and sacral sections are added (1 vertebra each);
in reptiles :
Cervical (8 vertebrae, I - atlas, II - axis);
Thoracic (articulated with the ribs, some of which are articulated with the sternum in front);
Lumbar, sacral (contain a larger number of vertebrae);
Tail;
in birds :
Cervical (up to 25 vertebrae);
Chest;
The lumbar sacrum and part of the caudal vertebrae are firmly connected, forming a powerful sacrum;
in mammals :
Cervical (7 vertebrae);
Thoracic (9-24 vertebrae);
Lumbar (3-9 vertebrae);
Sacral (vertebrae fuse together);
Caudal (coccygeal).
In humans, there are various defects in the formation of the spinal canal, through which spina bifida can emerge, as well as the formation of pathological curves, including lateral ones (scoliosis), and the caudal region can be preserved.
The head skeleton is divided into the cerebral and visceral skull. In the process of evolution, there was a decrease in the number of bones of the brain skull, replacement of cartilaginous parts with bone, and a decrease in the number of moving elements. The number of branchial arches of the visceral skeleton also progressively decreased. The first and second branchial arches have changed and given rise to the jaws, bones of the facial skull, auditory bones, and cartilage of the larynx.
In fish, arch I (maxillary, consists of the palatoquadrate and Meckel's cartilages) is directly connected to arch II (consists of hyomandibular cartilage and hyoid). This type of connection between the brain and facial skull is called hyostyle(since the II arch connects to the hyomandibular cartilage). Beginning with amphibians, the first arch, with its palatoquadrate cartilage, fuses with the base of the cranium. This type of connection is called autostyle. The auditory bone column is formed from the hyomandibular cartilage, from which the stapes is subsequently formed. With ontogeny disorders, a person may have only one of these auditory bones (as in amphibians and reptiles) instead of three.
Let us consider the structure of the hearing organ in various classes of vertebrates. U fish There is only an inner ear, which includes the membranous labyrinth with otoliths, semicircular canals and the cochlear rudiment.
U amphibians There is an inner ear of the same structure and a middle ear with one auditory bone and an eardrum.
U reptiles the inner ear consists of the cochlea and semicircular canals, the middle ear is the same as in amphibians, there is a rudiment of the external auditory canal.
U birds the external auditory canal is well formed.
U mammals the inner ear includes the cochlea 2.5 turns, semicircular canals, saccule, utricle; middle ear - the tympanic cavity in which the auditory bones are located - the stapes (a remnant of the hyomandibular cartilage), the incus (a remnant of the quadratus palatocartilage), the malleus (a remnant of Meckel's cartilage), there is a tympanic membrane, the external includes the external auditory canal and the auricle.
Thus, in the visceral skeleton, in the process of evolution, cartilaginous jaws were replaced by bone ones, the type of attachment of the jaws changed, and the function of the elements of the visceral arches changed.
In the skeleton of the limbs, belts with a movably attached skeleton of free limbs were formed; a five-fingered limb of the terrestrial type was formed; the number of bones in the distal sections decreased; the proximal parts of the limbs were lengthened and the distal parts were shortened.
A person may have the following defects: polydactyly And polyphalanx(increase in the number of fingers and phalanges, respectively); moving the belt of the upper limbs to the level of the I-II thoracic vertebrae - Sprengel's disease; congenital anomalies of the ribs and chest deformities - funnel-water, keeled and etc.
PHYLOGENESIS OF MUSCLES
Muscles perform the function of movement and are divided into:
Somatic, which develops from myotomes and is innervated by spinal nerves (striated);
Visceral - develops from other parts of the mesoderm, innervated by autonomic nerves, can be striated and smooth.
In lower chordates, the visceral muscles are located in the region of the gill arches, and gradually the muscles of mastication, the muscles of the pharynx, then the muscles of the neck and shoulder girdle, as well as facial muscles, are formed from it. The muscles of the neck and face have complex innervation, which is associated with the stages of phylogenesis. Somatic muscles in lower chordates are located in the form of myomeres, then the muscles of the limbs are formed, the shape, size and functions of the muscles become more diverse, their innervation increases, especially the free limbs in primates. The muscles of the trunk are reduced in the process of evolution.
The phylogeny of organ systems of chordates is considered in accordance with the progressive direction of evolution of this type of animals from the subphylum Cranial to the class Mammals. The organization of organ systems of the class Birds is not described due to the fact that birds evolved from reptiles much later than mammals and are a side branch of the evolution of chordates.
Outer coverings
Veils Any animal always performs the function of perceiving external irritations, and also protects the body from the harmful influences of the environment. The intensification of the first function of the integument leads, in the process of evolution of multicellular animals, to the emergence of a nervous system and sensory organs. The intensification of the second function is accompanied by differentiation. An expansion of functions is also characteristic, as a result of which the skin, as a protective organ, also participates in gas exchange, thermoregulation and excretion, and feeding of offspring. This is due to the complication of the structure of the skin layers, the appearance and further transformation of numerous appendages and glands.
In all chordates, the skin has a dual - ecto- and mesodermal - origin. The epidermis develops from the ectoderm, and the dermis from the mesoderm. Skullless are characterized by a weak degree of differentiation of both layers of skin. The epidermis is single-layered, cylindrical, containing single-celled mucous glands; the dermis is loose and contains a small number of connective tissue cells.
In the Vertebrate subtype, the epidermis becomes multilayered, with cells in the lower layer constantly multiplying, and in the upper layers they differentiate, die and exfoliate. Connective tissue fibers appear in the dermis, giving the integument strength. The skin forms appendages, diverse depending on the lifestyle and level of organization, as well as glands that perform various functions.
In fish, the glands in the epidermis are single-celled. Like the lancelet, they secrete mucus that facilitates movement in the water. The body of fish is covered with scales that have different structures depending on their systematic position. The scales of cartilaginous fish are called placoid. It has the shape of a spike and consists of dentin covered on the outside with enamel (Fig. 14.1). Dentin is of mesodermal origin; it is formed due to the functioning of connective tissue cells that protrude from the outside in the form of a papilla. Enamel, which is a harder non-cellular substance than dentin, is formed by the papilla of the epidermis and covers the outside of the placoid scales.
The entire surface of the body of cartilaginous fish, as well as the oral cavity, the mucous membrane of which comes from the ectoderm, is covered with placoid scales. Naturally, the functions of scales in the oral cavity are related to the capture and retention of food, so they are greatly enlarged and are teeth. Bony fish have a different type of scales. It looks like thin round bone plates covered with a thin layer of epidermis. The bone scale develops entirely at the expense of the dermis, but is related in origin to the primitive placoid scale.
The skin of primitive extinct amphibians - stegocephalians - corresponded to the skin of fish and was also covered with scales. Modern amphibians have thin, smooth skin without scales, which takes part in gas exchange. This is facilitated by the presence of a large number of multicellular mucous glands, the secret of which constantly moisturizes the skin and has bactericidal properties. Some skin glands of a number of amphibians differentiated into toxin-producing organs that protect them from enemies (see section 23.1).
Rice. 14.1. Laying of placoid scales:
1 - enamel-forming cells, 2- epidermis, 3- enamel, 4- dentin-forming scleroblasts, 5- dentine, 6- dermal papilla
Reptiles that have completely transitioned to a terrestrial existence have dry skin that does not participate in respiration. Upper layer of the epidermis keratinizes. The horny scales of some reptiles are thin and elastic, while in others they merge together, forming, like in turtles, a powerful horny shell. Most reptiles molt as they grow, periodically shedding their horny cover. Modern reptiles do not have skin glands.
The skin of mammals is built most complexly due to the variety of functions they perform. Various derivatives of the skin are characteristic: hair, claws, horns, hooves, as well as sweat, sebaceous and mammary glands. More primitive mammals - insectivores, rodents and some others - along with hair, also retained horny scales on the tail. Their hair grows in the spaces between the scales, in groups of 3-7. In more advanced mammals that have lost scales, the same arrangement of hair is retained (Fig. 14.2), covering almost the entire body, except for some areas, for example, the soles and palms of humans.
Hair many mammals are differentiated into typical ones, serving for thermoregulation, and large, or vibrissae, the bases of which are associated with sensory nerve endings. In most mammals, the vibrissae are located in the mouth and nose; in primates they are reduced due to the increased tactile function of the forelimbs; in many oviparous and marsupials they are scattered throughout the body. This fact may indicate that the hair of the ancestors of mammals primarily performed tactile functions, and then, as the amount of hair increased, it began to take part in thermoregulation. During human ontogenesis, a larger number of hair buds are formed, but by the end of embryogenesis, a reduction of most of them occurs.
Sweat glands mammals are homologous to the skin glands of amphibians. Their secretion can be mucous and contain proteins and fat. Some sweat glands differentiated in early mammals into mammary glands. In oviparous animals (platypus, echidna), the mammary glands are similar to the sweat glands in structure and development. Along the edges of the developing nipple of the mammary gland, successive transitions from typical sweat glands to mammary glands can be found (Fig. 14.3). The number of mammary glands and nipples correlates with fertility (from 25 to one pair), but in the embryogenesis of all mammals, “milky lines” are formed on the abdominal surface, stretching from the armpit to the groin. Subsequently, the nipples differentiate along these lines, most of which then undergo reduction and disappear. Thus, in human embryogenesis, five pairs of nipples are initially formed, and subsequently only one remains (Fig. 14.4).

Rice. 14.4. Embryogenesis of the human anterior abdominal wall. A - embryo at 5 weeks of age (milky lines visible); B - differentiation of five pairs of nipples; IN - embryo at 7 weeks of age

Rice. 14.5. Atavistic anomalies of skin development.
A - hypertrichosis; B - polymastia
Sebaceous glands are formed in the skin only in mammals. Their secretion, lubricating the hair and skin surface, gives it non-wetting and elasticity.
The ontogeny of the integument and appendages of the skin of mammals and humans reflects their evolution according to the type of archallaxis. Indeed, neither the rudiments of horny scales characteristic of reptiles, nor the earlier forms of skin appendages recapitulate in their embryogenesis. At the same time, at the stage of secondary organogenesis, the rudiments of hair follicles immediately develop. Disturbances in the early ontogenesis of human skin can cause the occurrence of some minor atavistic malformations: hypertrichosis (increased hair growth), polythelia (increased number of nipples), polymastia (increased number of mammary glands) (Fig. 14.5). All of them are associated with a violation of the reduction of the excess number of these structures and reflect the evolutionary connection of humans with the closest ancestral forms - mammals. This is why it is impossible for humans and other mammals to give birth to offspring with atavistic skin features characteristic of more distant ancestors. One of the most well-known signs of prematurity in newborns is increased skin hair growth. Shortly after birth, excess hair usually falls out and hair follicles become smaller.
MUSCULOCAL SYSTEM
Phylogenesis of motor function underlies the progressive evolution of animals. Therefore, the level of their organization primarily depends on the nature of motor activity, which is determined by the characteristics of the organization musculoskeletal system, undergone major evolutionary transformations in the phylum Chordata due to changes in habitats and changes in forms of locomotion. Indeed, the aquatic environment in animals that do not have an exoskeleton involves monotonous movements due to the bending of the entire body, while life on land is more conducive to their movement with the help of their limbs.
Let us consider separately the evolution of the skeleton and muscular system.
Skeleton
In chordates internal skeleton. According to its structure and functions, it is divided into the axial skeleton, the skeleton of the limbs and the head.
Axial skeleton
In the Skullless subtype there is only axial skeleton in the form of a chord. It is built of highly vacuolated cells, tightly adjacent to each other and covered on the outside with common elastic and fibrous membranes. The elasticity of the chord is given by the turgor pressure of its cells and the strength of the membranes. The notochord is formed in the ontogenesis of all chordates and in more highly organized animals performs not so much a supporting, but a morphogenetic function, being an organ that carries out embryonic induction.
Throughout life in vertebrates, the notochord is retained only in cyclostomes and some lower fish. In all other animals it is reduced. In humans, in the postembryonic period, rudiments of the chord are preserved in the form of nucleus pulposus intervertebral discs. Preservation of an excess amount of notochordal material when its reduction is impaired is fraught with the possibility of developing tumors in humans - chord, arising on its basis.
In all vertebrates, the notochord is gradually replaced vertebrae, developing from the sclerotomes of somites, and is functionally replaced spinal column. This is one of the clearest examples of homotopic organ substitution (see § 13.4). The formation of vertebrae in phylogeny begins with the development of their arches, covering the neural tube and becoming sites of muscle attachment. Beginning with cartilaginous fish, cartilagination of the shell of the notochord and the growth of the bases of the vertebral arches are detected, as a result of which the vertebral bodies are formed. The fusion of the upper vertebral arches above the neural tube forms the spinous processes and the spinal canal, which encloses the neural tube (Fig. 14.6).

Rice. 14.6. Vertebral development. A-early stage; B- next stage:
1 -chord, 2- chord shell, 3- upper and lower vertebral arches, 4- spinous process, 5- ossification zones, 6-notochord rudiment, 7 - vertebral cartilaginous body
Replacing the notochord with the spinal column - a more powerful support organ with a segmental structure - allows you to increase the overall size of the body and activates motor function. Further progressive changes in the spinal column are associated with tissue substitution - the replacement of cartilaginous tissue with bone, which is found in bony fish, as well as with its differentiation into sections.
Fish have only two parts of the spine: trunk And tail. This is due to their movement in water due to the bending of the body.
Amphibians also acquire cervical And sacral departments, each represented by one vertebra. The first provides greater mobility of the head, and the second provides support for the hind limbs.
In reptiles, the cervical spine lengthens, the first two vertebrae of which are movably connected to the skull and provide greater mobility of the head. Appears lumbar a section still weakly delimited from the thoracic one, and the sacrum already consists of two vertebrae.
Mammals are characterized by a stable number of vertebrae in the cervical region, equal to 7. Due to the great importance in the movement of the hind limbs, the sacrum is formed by 5-10 vertebrae. The lumbar and thoracic regions are clearly demarcated from each other.
In fish, all the trunk vertebrae bear ribs that are not fused with each other or with the sternum. They give the body a stable shape and provide support for the muscles that bend the body in the horizontal plane. This function of the ribs is preserved in all vertebrates that perform serpentine movements - in tailed amphibians and reptiles, therefore, their ribs are also located on all vertebrae except the caudal ones.
In reptiles, part of the thoracic ribs fuses with the sternum, forming the chest, and in mammals the chest includes 12-13 pairs of ribs.

Rice. 14.7. Developmental anomalies of the axial skeleton. A - vestigial cervical ribs (shown by arrows); B - Non-fusion of the spinous processes of the vertebrae in the thoracic and lumbar regions. Spina bifida
The ontogeny of the human axial skeleton recapitulates the main phylogenetic stages of its formation: during the period of neurulation, the notochord is formed, which is subsequently replaced by a cartilaginous and then a bone spine. A pair of ribs develops on the cervical, thoracic and lumbar vertebrae, after which the cervical and lumbar ribs are reduced, and the thoracic ribs fuse in front with each other and with the sternum, forming the rib cage.
Disruption of the ontogenesis of the axial skeleton in humans can be expressed in such atavistic developmental defects as nonfusion of the spinous processes of the vertebrae, resulting in the formation of spinabifida - spinal canal defect. In this case, the meninges often protrude through the defect and a spina bifida(Fig. 14.7).
At the age of 1.5-3 months. The human embryo has a caudal spine consisting of 8-11 vertebrae. Violation of their reduction subsequently explains the possibility of the occurrence of such a well-known anomaly of the axial skeleton as persistence of the tail.
Violation of the reduction of the cervical and lumbar ribs underlies their preservation in postnatal ontogenesis.
Head skeleton
The anterior continuation of the axial skeleton is axial, or brain, skull, serving to protect the brain and sensory organs. It develops next to it visceral, or facial skull, forming a support for the anterior part of the digestive tube. Both parts of the skull develop differently and from different rudiments. At the early stages of evolution and ontogenesis they are not connected with each other, but later this connection arises.

Rice. 14.8. Human skull with methodical suture (indicated by arrow)
In the posterior part of the axial skull during development, traces of segmentation are found, therefore it is believed that it is the result of the fusion of the anlage of the anterior vertebrae with each other. The composition of the brain skull also includes the anlage of cartilaginous capsules of mesenchymal origin surrounding the organs of hearing, smell and vision. In addition, the part of the brain skull (lying anterior to the sella turcica), which does not have segmentation, apparently develops as a neoplasm due to an increase in the size of the forebrain.
Phylogenetically, the brain skull went through three stages of development: membranous, cartilaginous And bone.
In cyclostomes it is almost entirely membranous and does not have an anterior, unsegmented part.
The skull of cartilaginous fish is almost entirely cartilaginous, and includes both the posterior, primarily segmented part, and the anterior one.
In bony fish and other vertebrates, the axial skull becomes bony due to the processes of ossification of cartilage in the area of its base (basal, sphenoid, ethmoid bones) and due to the appearance of integumentary bones in its upper part (parietal, frontal, nasal bones). The bones of the axial skull undergo oligomerization in the process of progressive evolution. The appearance of a large number of ossification zones and their subsequent fusion together during the formation of such bones as the frontal, temporal, etc., indicate this. Widely known in humans are such anomalies of the brain skull as the presence of interparietal bones, as well as two frontal bones with a metopic suture between them (Fig. 14.8). They are not accompanied by any pathological phenomena and are therefore usually discovered by chance after death.
The visceral skull also appears for the first time in lower vertebrates. It is formed from mesenchyme of ectodermal origin, which is grouped in the form of arch-shaped condensations in the spaces between the gill slits of the pharynx. The first two arches receive particularly strong development and give rise to the maxillary and hyoid arches of adult animals. The following arches, numbering 4-5 pairs, perform a supporting function for the gills and are called gills.
In cartilaginous fish, in front of the jaw arch there are usually 1-2 more pairs of premaxillary arches, which are rudimentary in nature. This indicates that the ancestors of vertebrates had a greater number of visceral arches than 6 or 7, and their differentiation occurred against the background of oligomerization.
The jaw arch consists of two cartilages. The top one is called palatoquadrate, he performs the function of the primary upper jaw. Lower, or Meckel, cartilage - primary lower jaw. On the ventral side of the pharynx, Meckel's cartilages are connected to each other in such a way that the jaw arch encloses the oral cavity in a ring. The second visceral arch on each side consists of hyomandibular cartilage fused to the base of the skull, and hyoid connected to Meckel's cartilage. Thus, in cartilaginous fishes, both primary jaws are connected to the axial skull through a second visceral arch, in which the hyomandibular cartilage acts as a suspension to the brain skull. This type of connection between the jaws and the axial skull is called hyostyle(Fig. 14.9).
In bony fishes, the replacement of primary jaws with secondary ones begins, consisting of false bones - the jaw and premaxilla on top and the tooth below. The palatoquadrate and Meckel's cartilages decrease in size and move posteriorly. The hyomandibular cartilage continues to function as a suspension, so the skull remains hyostylous.
Amphibians, in connection with the transition to terrestrial existence, have undergone significant changes in the visceral skull. The branchial arches are partially reduced, and partially, changing their functions, they become part of the cartilaginous apparatus of the larynx. The maxillary arch with its upper element - the palatine quadrate cartilage - fuses completely with the base of the brain skull, and the skull thus becomes autostyle. The hyomandibular cartilage, greatly reduced and freed from the function of suspension, located in the region of the first gill cleft inside the auditory capsule, took on the function of the auditory ossicle - a column - transmitting sound vibrations from the outer to the inner ear.
The visceral skull of reptiles is also autostyle. The jaw apparatus is characterized by a higher degree of ossification than that of amphibians. Part of the cartilaginous material of the gill arches is part of not only the larynx, but also the trachea.
The lower jaw of mammals articulates with the temporal bone with a complex joint that allows not only the capture of food, but also complex chewing movements.
One auditory ossicle - column,- characteristic of amphibians and reptiles, decreasing in size, turns into stapes, and the rudiments of the palatoquadrate and Meckel’s cartilages, completely leaving the jaw apparatus, are transformed, respectively, into anvil And hammer. Thus, a single functional chain of three auditory ossicles in the middle ear is created, characteristic only of mammals (Fig. 14.9).

Rice. 14.9. Evolution of the first two visceral branchial arches of vertebrates.
A- cartilaginous fish; B- amphibian; IN- reptile; G- mammal:
1 -palatoquadrate cartilage, 2-Meckel's cartilage, 3- hyomandibular cartilage, 4-hyoid, 5- column, 6- overlay bones of secondary jaws, 7-anvil, 8- stapes, 9- hammer; homologous formations are indicated by corresponding shading
Recapitulation of the main stages of phylogenesis of the visceral skull also occurs in human ontogenesis. Violation of the differentiation of the elements of the maxillary gill arch into the auditory ossicles is a mechanism for the formation of such a malformation of the middle ear as the location in the tympanic cavity of only one auditory ossicle - the column, which corresponds to the structure of the sound transmitting apparatus of amphibians and reptiles.
Limb skeleton
Chordates have unpaired and paired limbs. Unpaired (dorsal, caudal and anal fins) are the main organs of locomotion in amphibians, fish and, to a lesser extent, in caudate amphibians. Fish also develop paired limbs - pectoral and pelvic fins, on the basis of which paired limbs of terrestrial quadrupeds subsequently develop.
Let's take a closer look at the origin and evolution of paired limbs.
In fish larvae, as well as in modern skullless fish, lateral skin folds called metapleural(Fig. 14.10). They have neither a skeleton nor their own muscles, performing a passive role - stabilizing the body position and increasing the area of the abdominal surface, facilitating movement in the aquatic environment. Probably, in the ancestors of fish transitioning to a more active lifestyle, muscle elements and cartilaginous rays appeared in these folds, associated with somites in origin and therefore located metamerically. Such folds, having acquired mobility, can act as depth rudders, however, for changing the position of the body in space, their anterior and posterior sections are of greater importance, as they are the most distant from the center of gravity. Therefore, evolution followed the path of intensifying the functions of the outer parts and weakening the functions of the central parts.

Rice. 14.10. Formation of the fore and hind limbs from metapleural folds: I-III-hypothetical stages of evolution
As a result, pectoral fins developed from the anterior sections of the folds, and ventral fins from the posterior ones (Fig. 14.10). It is possible that the formation of only two pairs of limbs on the lateral sides of the body was preceded by the disintegration of continuous folds into a number of paired fins, of which the anterior and posterior ones were also more important. This is evidenced by the existence of fossil remains of the most ancient low-organized fish with numerous fins (Fig. 14.11). Due to the fusion of the bases of the cartilaginous rays, brachial And pelvic girdle. Rest their areas differentiated into skeleton of free limbs.

Rice. 14.11. An ancient shark-like fish with numerous paired limbs
In most fish, the skeleton of paired fins is divided into a proximal section, consisting of a small number of cartilaginous or bony plates, and a distal section, which includes a large number of radially segmented rays. The fins are connected to the girdles of the limbs inactively. They cannot serve as support for the body when moving along the bottom or land. In lobe-finned fish, the skeleton of paired limbs has a different structure. The total number of their bone elements is reduced, and they are larger in size. The proximal section consists of only one large bone element, corresponding to the humerus or femur of the forelimbs or hind limbs. This is followed by two smaller bones, homologous to the ulna and radius or tibia and fibula. 7-12 radially located rays rest on them. In connection with the girdles of the limbs of such a fin, only homologs of the humerus or femur are involved, therefore the fins of lobe-finned fish are actively mobile (Fig. 14.12, A, B) and can be used not only to change the direction of movement in water, but also to move along a solid substrate.
The life of these fish in shallow drying up reservoirs in the Devonian period contributed to the selection of forms with more developed and mobile limbs. The presence of additional respiratory organs in them (see Section 14.3.4) became the second prerequisite for reaching land and the emergence of other adaptations to terrestrial existence, which resulted in the origin of amphibians and the entire Tetrapoda group. Their first representatives - stegocephals - had seven- and five-fingered limbs that retained similarities with the fins of lobe-finned fish (Fig. 14.12, B)

Rice. 14.12. Skeleton of the limb of a lobe-finned fish ( A), its base ( B) and the skeleton of the front paw of Stegocephalus ( IN):I- humerus, 2-ulna, 3- radius
In the skeleton of the wrist, the correct radial arrangement of bone elements in 3-4 rows is preserved, in the metacarpus there are 7-5 bones, and then the phalanges of 7-5 fingers also lie radially.
In modern amphibians, the number of fingers in the limbs is five or they are oligomerized to four.
Further progressive transformation of the limbs is expressed in an increase in the degree of mobility of bone joints, in a decrease in the number of bones in the wrist, first to three rows in amphibians and then to two in reptiles and mammals. At the same time, the number of phalanges of the fingers also decreases. Lengthening of the proximal parts of the limb and shortening of the distal parts are also characteristic.
The arrangement of the limbs also changes during evolution. If in fish the pectoral fins are at the level of the first vertebra and are turned to the sides, then in terrestrial vertebrates, as a result of the complication of orientation in space, a neck appears and head mobility occurs, and in reptiles and especially mammals, due to the elevation of the body above the ground, the forelimbs move posteriorly and are oriented not horizontally, but vertically. The same applies to the hind limbs.
The variety of living conditions provided by the terrestrial way of life provides a variety of forms of movement: jumping, running, crawling, flying, digging, climbing rocks and trees, and when returning to the aquatic environment, swimming. Therefore, in terrestrial vertebrates one can find both an almost unlimited variety of limbs and their complete secondary reduction, and many similar limb adaptations in various environments have repeatedly arisen convergently (Fig. 14.13). However, during the process of ontogenesis, most terrestrial vertebrates exhibit common features in the development of limbs: the formation of their primordia in the form of poorly differentiated folds, the formation of six or seven digit primordia in the hand and foot, the outermost of which are soon reduced and only five subsequently develop (Fig. 14.14 ).

Rice. 14.13. Skeleton of the forelimb of terrestrial vertebrates. A-frog- B-salamander; IN-crocodile; G-bat; D-Human: 1 -humerus, 2-radius bone, 3 - wrist bones, 4 - metacarpus, 5 - phalanges of fingers, 6 -elbow bone
Rice. 14.14. Structure of a developing vertebrate limb: pp - prepollex, pin - postminimus - additional rudimentary digits I and VII
It is interesting that in the embryogenesis of higher vertebrates, not only the structure of the limbs of the ancestors recapitulates, but also the process of their heterotopy. Thus, in humans, the upper limbs are formed at the level of the 3rd-4th cervical vertebrae, and the lower ones - at the level of the lumbar vertebrae. At the same time, the limbs receive innervation from the corresponding parts of the spinal cord. Heterotopia of the limbs is accompanied by the formation of the cervical, lumbar and sacral nerve plexuses, the nerves of which are connected, on the one hand, to those segments of the spinal cord from which they grew at the time of the formation of the limbs, and on the other, to the limbs that have moved to a new place (Fig. 14.15; see .see also section 14.2.2.2).
Numerous disorders are possible in human ontogenesis, leading to the formation of atavistic congenital malformations of the limbs. So, polydactyly, or an increase in the number of fingers, inherited as an autosomal dominant trait, is the result of the development of the anlages of additional fingers, which are normally characteristic of distant ancestral forms. The phenomenon of polyphalanx is known, characterized by an increase in the number of phalanges, usually the thumb. Its occurrence is based on the development of three phalanges in the first digit, as is normally observed in reptiles and amphibians with undifferentiated digits. Bilateral polyphalanx is inherited in an autosomal dominant manner.
A serious malformation is a violation of heterotopia of the upper limb girdle from the cervical region to the level of the 1st-2nd thoracic vertebrae. This anomaly is called Sprengel's disease or congenital high position of the scapula (Fig. 14.16). It is expressed in the fact that the shoulder girdle on one or both sides is several centimeters higher than the normal position. Due to the fact that such a disorder is often accompanied by anomalies of the ribs, thoracic spine and deformation of the shoulder blades, one should think that the mechanisms of its occurrence are not only a violation of the movement of organs, but also a resulting violation of morphogenetic correlations (see § 13.4).
A comparative anatomical review of the evolution of the chordate skeleton indicates that the human skeleton is completely homologous to the supporting apparatus of ancestral and related forms. Therefore, many malformations of its development in humans can be explained by the relationship of mammals with reptiles, amphibians and fish. However, in the process of anthropogenesis, skeletal features appeared that are characteristic only of humans and are associated with their upright posture and work activity. These include: 1) changes in the foot that has ceased to perform a grasping function, expressed in the loss of the ability to oppose the big toe and the appearance of its arches, which serve for shock absorption when walking; 2) changes in the spinal column - its S-shaped bend, providing plasticity of movements in a vertical position; 3) changes in the skull - a sharp decrease in its facial part and an increase in the brain, anterior displacement of the foramen magnum, an increase in the mastoid process and smoothing of the occipital relief, to which the neck muscles and nuchal ligament are attached; 4) specialization of the upper (fore) limbs as an organ of labor; 5) the appearance of a chin protrusion in connection with the development of articulate speech.

Rice. 14.15. Formation of the forelimbs, their heterotopia and innervation in human ontogenesis. A-ingrowth of cervical myotomes into the developing forelimb of the embryo; B-development of skin innervation of the hand; IN- location of the cervical and brachial plexuses involved in the innervation of the arm:
1 -cervical myotomes, 2- thoracic myotomes, 3 -lumbar myotomes; the letters C, T, L indicate the cervical, thoracic and lumbar segments

Rice. 14.16. Sprengel's disease (see text for explanation)
Despite the fact that the formation of anatomical and morphological features of the human skeleton is apparently complete, adaptations to upright walking in humans are, like all adaptations in general, of a relative nature. Thus, with heavy physical activity, displacement of the vertebrae or intervertebral discs is possible. Having switched to upright walking, man has lost the ability to run quickly and moves much slower than most four-legged animals.
Naturally, during intrauterine development, the skeletal features that characterize a person as a unique biological species are formed at its final stages or even, such as the S-shaped spine, in the early postnatal period of development. They are actually anabolics that arose during the phylogenesis of primates. That's why atavistic anomalies skeleton associated with delays in the development of characteristics characteristic only of humans are the most common. They practically do not reduce vitality, but children with them need orthopedic correction, gymnastics and massage. Such anomalies include mild forms of congenital flat feet, club feet, a narrow chest, absence of a chin protrusion and some others.
Muscular system
In representatives of the phylum Chordata, the muscles are divided according to the nature of development and innervation into somatic and visceral.
Somatic musculature develops from myotomes and is innervated by nerves, the fibers of which exit the spinal cord as part of the ventral roots of the spinal nerves. Visceral muscles develops from other parts of the mesoderm and is innervated by the nerves of the autonomic nervous system. All somatic muscles are striated, and visceral muscles can be either striated or smooth (Fig. 14.17).

Rice. 14.17. Somatic and visceral muscles of vertebrates:
1 -somatic muscles developing from myotomes, 2- visceral muscles of the gill region
Visceral muscles
The most significant changes have undergone the visceral muscles associated with the visceral arches of the anterior part of the digestive tube. In lower vertebrates, most of this muscle is represented by the general constrictor of the visceral apparatus - m. constrictor superficialis, covering the entire area of the gill arches on all sides. In the area of the jaw arch this muscle is innervated trigeminal nerve(V), in the area of the hyoid arch - facial(VII), in the area of the first gill arch - glossopharyngeal(IX), finally, its part lying more caudally - wandering nerve (X). In this regard, all derivatives of the corresponding visceral arches and the muscles associated with them are subsequently innervated in all vertebrates by the listed nerves.
In the anterior part of the compressor there is a large muscular mass that serves the jaw apparatus. Behind the visceral apparatus, the trapezius muscle m is differentiated. trapezius, attached in separate bundles to the last gill slits and the anterior edge of the dorsal part of the shoulder girdle. Part of the superficial constrictor in the area of the hyoid arch in reptiles grows, covers the neck from below and from the sides and forms the cervical constrictor m. sphincter colli. In mammals, this muscle is divided into two layers: deep and superficial. The deep one retains the same name, and the superficial one is called platysma myoides and is located subcutaneously. These two muscles grow over the entire head region and give rise to a complex system of facial subcutaneous muscles, which in primates and humans is called mimic. Therefore, all mimic
LECTURE MYOLOGY PHYLOGENESIS, ONTOGENESIS AND FUNCTIONAL ANATOMY OF THE MUSCULAR SYSTEM Performed by: Vladimirova Ya. B. Kokoreva T. V.

 Muscles or muscles (from Latin musculus - mouse, small mouse) are organs of the body of animals and humans, consisting of elastic, elastic muscle tissue, capable of contracting under the influence of nerve impulses. Designed to perform various actions: body movements, contraction of vocal cords, breathing. Muscles consist of 86.3% water. There are 640 muscles in the human body
Muscles or muscles (from Latin musculus - mouse, small mouse) are organs of the body of animals and humans, consisting of elastic, elastic muscle tissue, capable of contracting under the influence of nerve impulses. Designed to perform various actions: body movements, contraction of vocal cords, breathing. Muscles consist of 86.3% water. There are 640 muscles in the human body
 Motivation: - - - possibilities of the movement performed, volume of movement; active or passive movements are triggered by one or another muscle group; by influencing the muscular system, we change the general condition; muscle relief is a guide for the topography of blood vessels and nerves; muscle transplantation, that is, the muscle can be “relearned”.
Motivation: - - - possibilities of the movement performed, volume of movement; active or passive movements are triggered by one or another muscle group; by influencing the muscular system, we change the general condition; muscle relief is a guide for the topography of blood vessels and nerves; muscle transplantation, that is, the muscle can be “relearned”.
 Development of muscles of cranial origin - from the head myotomes (sclerotomes) and mesenchyme of the branchial arches. Innervated by branches of cranial nerves of spinal origin - from the myotomes of the trunk of the embryo: from the ventral myotomes they are innervated by the anterior branches of the SMN; - from the dorsal myotomes they are innervated by the posterior branches of the SMN - Autochthonous muscles - muscles that remain at the site of their primary anlage. Truncofugal muscles are muscles that have moved from the trunk to the limbs. Truncopetal muscles are muscles that have moved from the limbs to the torso.
Development of muscles of cranial origin - from the head myotomes (sclerotomes) and mesenchyme of the branchial arches. Innervated by branches of cranial nerves of spinal origin - from the myotomes of the trunk of the embryo: from the ventral myotomes they are innervated by the anterior branches of the SMN; - from the dorsal myotomes they are innervated by the posterior branches of the SMN - Autochthonous muscles - muscles that remain at the site of their primary anlage. Truncofugal muscles are muscles that have moved from the trunk to the limbs. Truncopetal muscles are muscles that have moved from the limbs to the torso.



 Striated Smooth 1. The unit of organization is the myocyte. Length about 50 microns. Width from 6 microns. 2. Involuntary contraction Control by the autonomic nervous system The movement is wave-like, works slowly, since the nerve fiber does NOT approach every cell They come into action slowly, but persist for a long time Does not have an exact spatial orientation of cells 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiac The unit of organization is the muscle fiber - a set of myoblasts floating in the common cytoplasm (sarcoplasm). They have a common sarcolemma. Length about 40 -100 mm. Width from 7 mm. Voluntary contraction Control by the somatic nervous system Fast contraction, quick reaction, so each muscle fiber has a neuromuscular synapse Switch on quickly, but have a short-term effect Clear orientation of muscle fibers
Striated Smooth 1. The unit of organization is the myocyte. Length about 50 microns. Width from 6 microns. 2. Involuntary contraction Control by the autonomic nervous system The movement is wave-like, works slowly, since the nerve fiber does NOT approach every cell They come into action slowly, but persist for a long time Does not have an exact spatial orientation of cells 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiac The unit of organization is the muscle fiber - a set of myoblasts floating in the common cytoplasm (sarcoplasm). They have a common sarcolemma. Length about 40 -100 mm. Width from 7 mm. Voluntary contraction Control by the somatic nervous system Fast contraction, quick reaction, so each muscle fiber has a neuromuscular synapse Switch on quickly, but have a short-term effect Clear orientation of muscle fibers

 Between the muscle fibers there are thin layers of loose fibrous connective tissue - endomysium. The collagen fibers of the outer layer of the basement membrane are woven into it, which helps to combine forces during contraction of the myosymplasts. Thicker layers of loose connective tissue surround several muscle fibers, forming the perimysium and dividing the muscle into bundles. Several bundles are combined into larger groups, separated by thicker connective tissue layers. The connective tissue surrounding the surface of the muscle is called the epimysium.
Between the muscle fibers there are thin layers of loose fibrous connective tissue - endomysium. The collagen fibers of the outer layer of the basement membrane are woven into it, which helps to combine forces during contraction of the myosymplasts. Thicker layers of loose connective tissue surround several muscle fibers, forming the perimysium and dividing the muscle into bundles. Several bundles are combined into larger groups, separated by thicker connective tissue layers. The connective tissue surrounding the surface of the muscle is called the epimysium.
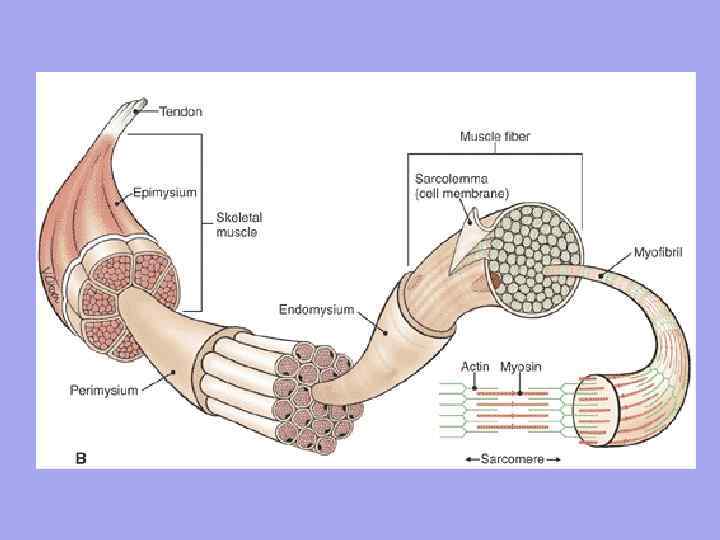
 Muscle as an organ contains connective tissue. Endomysium is a thin connective tissue that surrounds each muscle fiber and small groups of fibers. Perimysium – covers larger complexes of muscle fibers and muscle bundles.
Muscle as an organ contains connective tissue. Endomysium is a thin connective tissue that surrounds each muscle fiber and small groups of fibers. Perimysium – covers larger complexes of muscle fibers and muscle bundles.
 Significance of endomysium and perimysium 1. Through the endomysium and perimysium, vessels and nerves approach the muscle fiber. They form the stroma of the organ; 2. Muscle fibers are formed into bundles, bundles into muscles; 3. Since the endomysium is fused with the sarcolemma of the muscle fiber, therefore, the contracting muscle fiber can only stretch to a certain limit
Significance of endomysium and perimysium 1. Through the endomysium and perimysium, vessels and nerves approach the muscle fiber. They form the stroma of the organ; 2. Muscle fibers are formed into bundles, bundles into muscles; 3. Since the endomysium is fused with the sarcolemma of the muscle fiber, therefore, the contracting muscle fiber can only stretch to a certain limit
 The myofibrils in the fiber are surrounded by a shell - sarcolemma, and immersed in a special medium - sarcoplasm. Depending on the pigment and oxygen content, the fibers are divided into white and red. White fibers are anaerobic, contain more myofibrils and less sarcoplasm. They start up quickly, but cannot work for a long time. Example: sternocleidomastoid, gastrocnemius muscles. Red fibers are thick fibers. There is a lot of myoglobin in the sarcoplasm and cytochrome in the mitochondria, but fewer myofibrils. Slow to start, but last a long time. Example: back muscles, diaphragm.
The myofibrils in the fiber are surrounded by a shell - sarcolemma, and immersed in a special medium - sarcoplasm. Depending on the pigment and oxygen content, the fibers are divided into white and red. White fibers are anaerobic, contain more myofibrils and less sarcoplasm. They start up quickly, but cannot work for a long time. Example: sternocleidomastoid, gastrocnemius muscles. Red fibers are thick fibers. There is a lot of myoglobin in the sarcoplasm and cytochrome in the mitochondria, but fewer myofibrils. Slow to start, but last a long time. Example: back muscles, diaphragm.
 Each muscle has a network of blood vessels. Muscle contractions promote blood flow. In a relaxed, non-working muscle, most of the blood capillaries are closed to blood flow. When a muscle contracts, all blood capillaries immediately open.
Each muscle has a network of blood vessels. Muscle contractions promote blood flow. In a relaxed, non-working muscle, most of the blood capillaries are closed to blood flow. When a muscle contracts, all blood capillaries immediately open.

 The structure of a muscle Each muscle is connected at one end to one bone (the origin of the muscle), and at the other end to the other (the attachment of the muscle). The muscle is divided into: head, abdomen, and tail.
The structure of a muscle Each muscle is connected at one end to one bone (the origin of the muscle), and at the other end to the other (the attachment of the muscle). The muscle is divided into: head, abdomen, and tail.
 Motor nerve fibers approach each muscle fiber and sensory nerve fibers depart. The number of nerve endings in a muscle depends on the degree of functional activity of the muscles.
Motor nerve fibers approach each muscle fiber and sensory nerve fibers depart. The number of nerve endings in a muscle depends on the degree of functional activity of the muscles.

 Each muscle fiber is innervated independently and is surrounded by a network of hemocapillaries, forming a complex called a myon. A group of muscle fibers innervated by one motor neuron is called a motor unit. It is characteristic that muscle fibers belonging to one motor unit do not lie side by side, but are located mosaically among fibers belonging to other units.
Each muscle fiber is innervated independently and is surrounded by a network of hemocapillaries, forming a complex called a myon. A group of muscle fibers innervated by one motor neuron is called a motor unit. It is characteristic that muscle fibers belonging to one motor unit do not lie side by side, but are located mosaically among fibers belonging to other units.


 A tendon is a dense fibrous connective tissue cord that connects a muscle to or attaches to the skeleton.
A tendon is a dense fibrous connective tissue cord that connects a muscle to or attaches to the skeleton.
 peritenonium type IV collagen fibers endotenonium Collagen fibers of the tendon, intertwined with collagen fibers of the periosteum, are woven into the ground substance of the bone tissue, forming ridges, tubercles, tubercles, depressions, and depressions on the bones.
peritenonium type IV collagen fibers endotenonium Collagen fibers of the tendon, intertwined with collagen fibers of the periosteum, are woven into the ground substance of the bone tissue, forming ridges, tubercles, tubercles, depressions, and depressions on the bones.
 Fascia is connective tissue collagen fibers with a small admixture of elastic fibers Superficial temporal fascia Deep fascia of the thigh
Fascia is connective tissue collagen fibers with a small admixture of elastic fibers Superficial temporal fascia Deep fascia of the thigh
 1. 2. 3. 4. 5. Fascia separates the muscles from the skin and eliminates the displacement of the skin during the movements of contracting muscles. Fascia conserves the force of muscle contraction by eliminating friction between muscles during contraction. Fascia stretches large veins under tension, as a result of which blood from the periphery is “sucked” into these veins. Fascia is important as barriers that prevent the spread of infection and tumors. During operations, fascia helps determine the location of muscles, blood vessels, and viscera.
1. 2. 3. 4. 5. Fascia separates the muscles from the skin and eliminates the displacement of the skin during the movements of contracting muscles. Fascia conserves the force of muscle contraction by eliminating friction between muscles during contraction. Fascia stretches large veins under tension, as a result of which blood from the periphery is “sucked” into these veins. Fascia is important as barriers that prevent the spread of infection and tumors. During operations, fascia helps determine the location of muscles, blood vessels, and viscera.
 Classification of muscles Skeletal muscles vary in shape, structure, position relative to the axes of the joints, etc., and therefore are classified differently.
Classification of muscles Skeletal muscles vary in shape, structure, position relative to the axes of the joints, etc., and therefore are classified differently.

 III. According to the functional features, Static (strong) - short belly and long tendon. The muscles work with greater force, but with a smaller range of motion. Dynamic (dexterous) – long muscle bundles, short tendons. Muscles work with less force, but produce larger movements
III. According to the functional features, Static (strong) - short belly and long tendon. The muscles work with greater force, but with a smaller range of motion. Dynamic (dexterous) – long muscle bundles, short tendons. Muscles work with less force, but produce larger movements
 Accessory apparatus of muscles Skeletal muscles have an accessory apparatus that facilitates their functioning. n n n Fascia; Osteofascial sheaths; Synovial bursae; Synovial tendon sheaths; Muscle blocks; Sesamoid bones.
Accessory apparatus of muscles Skeletal muscles have an accessory apparatus that facilitates their functioning. n n n Fascia; Osteofascial sheaths; Synovial bursae; Synovial tendon sheaths; Muscle blocks; Sesamoid bones.
 Anomalies of muscle development are very common and are divided into three groups: 1. Absence of any muscle; 2. The presence of an additional muscle that does not exist in nature. 3. Additional bundles of existing muscle.
Anomalies of muscle development are very common and are divided into three groups: 1. Absence of any muscle; 2. The presence of an additional muscle that does not exist in nature. 3. Additional bundles of existing muscle.
 Developmental defects: Underdevelopment of the sternocleidomastoid muscle - Torticollis; Underdevelopment of the diaphragm. Cause of diaphragmatic hernia. Underdevelopment of the deltoid and trapezius muscles – Deformation of the shoulder girdle and shoulder
Developmental defects: Underdevelopment of the sternocleidomastoid muscle - Torticollis; Underdevelopment of the diaphragm. Cause of diaphragmatic hernia. Underdevelopment of the deltoid and trapezius muscles – Deformation of the shoulder girdle and shoulder


 I. Shape: Fusiform; Ribbon-shaped; Flat wide; Serrated; Long; n n n Square; Triangular; Round; Deltoid; Soleus, etc.
I. Shape: Fusiform; Ribbon-shaped; Flat wide; Serrated; Long; n n n Square; Triangular; Round; Deltoid; Soleus, etc.



 II. In the direction of muscle fibers With straight parallel fibers; With transverse ones; With circular; Pinnate: A. Unipinnate; Bipinnate; C. Multipinnate. B.
II. In the direction of muscle fibers With straight parallel fibers; With transverse ones; With circular; Pinnate: A. Unipinnate; Bipinnate; C. Multipinnate. B.

 IV. By function: Adductors; Diverters; Bending; Extensor; Pronators; n n Arch supports; Straining; Muscles are synergists; Muscles are antagonists.
IV. By function: Adductors; Diverters; Bending; Extensor; Pronators; n n Arch supports; Straining; Muscles are synergists; Muscles are antagonists.
 V. In relation to the joint: Single-joint; Two-joint; Multi-joint.
V. In relation to the joint: Single-joint; Two-joint; Multi-joint.
The muscular system develops in parallel with the development of the nervous system as its executive organ. In phylogenesis, it appears on the basis of the contractility properties of cell elements of the simplest multicellular organisms that respond to irritation. Initially, smooth muscle tissue appears in the animal’s body, still well preserving its cellular structure and connected to the nervous system through two neurons. (Efferent communication with the centers of the nervous system is carried out through a peripheral neuron.) With the improvement of the forms of translational movement and the associated supporting elements of the body - the skeleton, skeletal (somatic) muscle tissue appears, which already has a direct efferent connection with the central nervous system.
If in coelenterate animals the muscle elements are not yet isolated, but are represented by special muscle processes of epithelial cells, which differentiate into longitudinally located (in the ectoderm) and ring-shaped (in the endoderm), then in worms they are isolated, are no longer associated with the epithelium and form a musculocutaneous bag.
In chordates, more mobile animals, all muscles are divided into somatic and visceral. In adults, the somatic muscles are striated. They are represented by a longitudinal layer of paired lateral muscles, divided by membranes into separate metameres (segments). The lancelet still has smooth muscle tissue in its vessels and intestines. In vertebrates, like in the lancelet, muscles are divided into somatic and smooth, differing not only in structure, but also in the nature of the nerve connections.
The more complex the organism of vertebrates becomes in phylogenesis, the more skeletal muscles lose their metamerism, the more differentiated they become, dividing first into dorsal and ventral layers, and then into separate layers, sections, muscle groups and muscles.
In terrestrial vertebrates, all skeletal muscles are already divided into groups: muscles of the trunk, head and limbs. Smooth muscle tissue still remains mainly in the walls of blood vessels and internal organs. No matter how complex skeletal muscle is as an organ, it is connected to certain segments of the brain.
This connection is established from the first moments of the appearance of muscle elements and is not broken during phylogenetic transformations. In this regard, it was established that a group of muscle fibers receiving nerve endings (nerve plaques) from one nerve cell is a “muscle unit” . From which segment the muscle develops in the embryo, from the same segment it is innervated in the future. If a muscle moves during development, a nerve follows (a prime example of this is the diaphragm located behind the thoracic cord). If the muscle differentiates into separate parts, the nerve undergoes the same differentiation; if the muscle fuses, the nerves also fuse. Reduction of the muscle leads to reduction of the nerve connection. Such an invariable connection makes it possible to identify the paths of muscle development in embryogenesis and phylogenesis, and makes it possible to establish through which segments of the embryo it was formed.
Figure 91. Diagram of the external muscles of vertebrates
During the process of individual development (in ontogeny), muscles develop from several sources of mesoderm; 1) due to the material of the mesoderm myotomes; 2) from unsegmented mesoderm in the gill area. Therefore, muscles that develop due to the mesenchyme of the gill apparatus are called visceral. Initially, in the embryo, the anlage of muscle elements, as in phylogenesis, is segmented and differentiated into separate metameres, separated by connective tissue partitions.
In ontogenesis, the main stages are repeated, especially in the early period of phylogenesis (Fig. 91). Initially, the lateral muscle is laid along the body as a longitudinal paired cord, segmented by connective tissue septa into metamerically located myotomes, and each myotome receives the nerve of a certain nerve segment. Then a longitudinal connective tissue septum appears, dividing the myotome into dorsal and ventral parts. Subsequently, in place of this longitudinal septum, transverse processes of the vertebrae and ribs develop. In the same sequence, the division of nerve branches occurs, maintaining connections with the same nerve segments. Due to the material of the dorsal parts of the myotomes, the muscles of the spinal column subsequently develop, and due to the ventral parts, the muscles of the lateral thoracic and abdominal walls develop.
Somewhat later, muscle bundles grow into the limb buds from the dorsal and ventral sections of the myotomes (from five myotomes in the pectoral limbs and from eight in the pelvic limbs). With the differentiation of muscle layers into individual muscles, skeletal muscles become even more complex, especially with the separation of the muscles of the limbs, which are also innervated mainly by the ventral branches of the spinal nerves.
The muscles of the head and some muscles of the neck develop from the unsegmented mesoderm of the region of the first five branchial arches. From the rudiments of the first branchial arch, masticatory muscles associated with the V pair of cranial nerves develop, from the second - facial muscles associated with the VII pair of cranial nerves.
Muscle tissue, due to its ability to contract during the process of phylogenesis, was formed into separate organs - muscles - belt-shaped engines with strength capable of lifting heavy weights, supporting body weight on the legs, delivering blows, developing high speed when moving, etc.
Muscles, when contracting, not only cause movement (perform dynamic work), but also provide muscle tone, strengthening joints at a certain angle of combination with a stationary body, maintaining a certain posture (perform static work). Static work, especially in large (heavy) ungulates, is extremely difficult.
A certain muscle tone also performs an anti-gravity function. Together with tendons, fascia and ligaments of the skeleton, muscles have elastic properties, thanks to which they are reliable shock absorbers, giving the limbs greater springiness, and also work as micropumps that promote the removal of blood and lymph from organs (N. I. Ariinchin, 1987).
In each species of animal, in the process of evolution, the muscles, developing, determine their limits of strength, speed of movement, endurance, as well as mass, which is directly dependent on the species, age, gender, breed of the animal and their training. Muscle mass is approximately 42-47% of total body weight in cattle and horses, 34% in sheep, and 31% in pigs. Only work (training) of muscles helps to increase their mass both by increasing the diameter of muscle fibers (hypertrophy) and by increasing their number (hyperplasia).
The work of muscles is closely connected with the organ of balance and, to a large extent, with other sense organs. Thanks to this connection, the muscles provide balance to the body, precision of movements, and strength. Muscles are a powerful generator of the transformation of the chemical energy of food into thermal energy, the energy of elastic deformations into mechanical energy, which is used to move blood through the vessels and excite afferent impulses sent to the brain, as well as the transition into thermal energy.
Ten of Swords Tarot in layouts - meaning and interpretation
How long does it take to cook cherry jam with pits?
Professional qualities of a kindergarten speech therapist teacher
Modern lesson in the context of the federal state educational standard of the new generation
Lecture on biology on the topic "Phylogenesis of the musculoskeletal and nervous systems" Phylogenesis of the muscular system: patterns of development