Úkolystudovat tuto sekci:
- Tato část pomáhá pochopit způsoby evoluce organického světa, postupné komplikace nejen strukturální, ale také fyziologické a biochemické mechanismy, které vedly ke vzniku moderních kraniálních strunatců, včetně primátů a člověka;
- v procesu srovnávání je možné jednoznačně ověřit fyziologickou podmíněnost vzniku nebo komplikace určitých struktur, jakož i jejich vymizení, tj. lépe porozumět stavbě lidského těla;
- pomáhá porozumět lidské embryologii, protože podle základního biogenetického zákona formulovaného v roce 1866 Haeckelem a Müllerem: ontogeneze je krátké a rychlé opakování fylogeneze.
Proto se v embryogenezi lidé vyznačují výskytem struktur, funkcí a biochemických reakcí, které byly přítomny u jejich předchůdců strunatců. Takovým opakováním se říká rekapitulace. V ontogenezi člověka se většinou neobjevují struktury dospělých ryb, obojživelníků a plazů, ale pouze jejich larvy. Navíc rané fáze vývoje rekapitulují úplněji než ty pozdější.
Evoluční vývoj ale není prostým opakováním strukturálních rysů předků, ale předpokládá změny při formování orgánů a systémů . Tento:
- cenogeneze - adaptace embrya na jiné prostředí, které se u dospělých forem ztrácí, např. žábry u pulce; provizorní orgány amniot;
- fylembryogeneze - vývojové změny, které mají adaptivní význam:
A) anabolie (prodloužení) - přidání dalších stupňů, například svalové bránice savců, peří ptáků;
b) odchylka - odchylka od programu ve středních fázích morfogeneze, například tvorba sluchových kůstek, výskyt alveolární struktury plic místo buněčné; biogenetický zákon je v těchto případech částečně naplněn;
PROTI) archalaxa - odchylka ontogeneze hned na začátku, kdy není naplněn biogenetický zákon, např. tvorba ochlupení začíná ztluštěním epidermis, která je ponořena do koria;
G) heterochronie - změna doby tvorby struktury, například časná tvorba placentární amnionu; A heterotopie - změna umístění struktury, například vytvoření lidského ramenního pletence na úrovni krčních obratlů III-IV, který se poté přesune na úroveň I-II hrudních obratlů.
Při poruchách embryogeneze se u dospělých mohou vyvinout příznaky, které byly přítomny u jejich vzdálených předků - atavismy. Pokud snižují životaschopnost, jsou tzv atavistický(nebo rodový z anĉetre) neřesti. Atavismy vznikají kvůli:
Nedostatečný rozvoj orgánů (hypoplazie bránice, rozštěp patra - „rozštěp patra“ atd.);
Perzistence (zachování) embryonálních struktur (patent ductus ductus, laterální krční píštěle, umbilikální píštěle atd.);
Poruchy pohybu orgánů v ontogenezi (pánevní umístění orgánů, kryptorchismus atd.).
Uvažujme umístění strunatců ve světě zvířat a jejich klasifikace.
Kmen Chordata sjednocuje mnohobuněčné deuterostomy různých úrovní organizace. Navíc absolutně všichni strunatci, alespoň v jedné fázi vývoje, mají tři vlastnosti, které je odlišují od ostatních zvířat:
- osový komplex orgánů, který je založen na vnitřní kostře - notochordu, nad ním - nervové trubici, pod ní - trávicí trubici;
- do hltanu prostupují hltanové štěrbiny, které jsou u vyšších strunatců podepřeny útrobními oblouky;
- ocasní část
Kromě toho lze zaznamenat přítomnost žilního systému portální žíly (krev z gastrointestinálního traktu prochází játry) a strukturu pokožky epidermis a dermis, ale vrstva ostnokožců má podobnou strukturu.
Kmen Chordata zahrnuje čtyři podfyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Larvální strunatci (Urochordata) - ascidiánové;
- III. Skullless (Acrania) - lancelety;
- IV. Obratlovci (Vertebrata), které zahrnují několik tříd:
Skupina Anamnie (nižší obratlovci)
- Cyklostomy (Cyclostomata) - mihule, hagfish - nemají čelisti;
- supertřída Ryby:
A. třída Chrupavčité a pancéřové ryby (Chondrichthyes) - žraloci, rejnoci, chiméry;
b. třída Bony fish (Osteichthyes) - ostatní ryby;
- Obojživelníci (Amphibia):
A. Ocasáči - čolci, mloci;
b. Bezocasá zvířata - žáby, ropuchy, rosničky;
PROTI. Beznohé jsou ceciliany;
Skupina Amniota (vyšší obratlovci)
- Plazi (Reptilia):
A. Šupinatý - ještěrky, hadi;
b. Želvy;
PROTI. Krokodýli;
g. Beakheads - hatteria;
- Ptáci (Aves) - vedlejší větev evoluce, mají mnoho specifických adaptací
A. Běžci - pštrosi;
b. tučňáci;
PROTI. Keeleds - ostatní ptáci;
- Savci (Mammalia):
A. Oviparous - ptakopysk, echidna;
b. Vačnatci - klokani;
PROTI. Placenty - ostatní zvířata a lidé.
FYLOGENEZE VNĚJŠÍ INTEGRACE
Hlavní funkce kožní vrstvy jsou vymezení a ochrana mnohobuněčného organismu. Slupka obratlovců získává řadu dalších funkcí. U strunatců se krycí vrstva skládá z:
Epidermis vyvíjející se z ektodermu;
Dermis vyvíjející se z mezodermu.
Směry evoluce jsou následující:
Jednovrstvá epidermis (lanceleta) se stává vícevrstevnou (u obratlovců), vrstvy se diferencují a navenek keratinizují; objevují se deriváty epidermis - zrohovatělé šupiny, drápy, vlasy atd.
Tenká, želatinová pojivová tkáň dermis se zahušťuje a získává na síle v důsledku zvýšení počtu vláken.
Pokožka a dermis mají četné deriváty: chromatofory, šupiny, zrohovatělé štítky, lastury, destičky, zobák, drápy, peří, vlasy, rohy, kopyta. Všechny strunatce mají žlázy. U nižších živočichů jsou to jednobuněčné slizniční žlázy, počínaje rybami se objevují jedovaté žlázy, dále mazové žlázy (u ptáků), u savců se přidávají mléčné a potní žlázy.
Osoba může mít následující poruchy ontogeneze kožního povlaku: albinismus(plné) a vitiligo(neúplná) depigmentace kůže, alopecie(nepřítomnost) a hypertrichóza(nadměrný růst) vlasů, hyperkeratóza A pachyonychie- ztluštění epidermis a nehtů, resp. polythelia A polymastie- zvýšení počtu bradavek a mléčných žláz, resp.
FYLOGENEZE KOSTEL
Nižší strunatci žijí ve vodě, zatímco vyšší strunatci žijí převážně na souši. Přechod z hustšího vodního prostředí do vzdušného vedl k výrazné komplikaci kostry a svalů, zejména kostra končetin a jejich pletenců se stala složitější a zvýšil se počet specializovaných svalů obsluhujících končetiny. Hlavní funkce kostry: podpora, základ pro svalový úpon, účast na minerálním metabolismu.
Na notochordu je založena osová kostra, kolem notochordu se pak vytvořily obratle, ty se diferencovaly, zvětšil se počet částí páteře, u člověka se vlivem vzpřímeného držení těla vytvořily ohyby - 2 kyfózy a 2 lordózy a zbytky v. notochord se zachoval pouze v centrálních úsecích meziobratlových plotének.
Tím pádem, v rybách Existují dvě části páteře:
Trup (trmenové obratle kloubově spojené s žebry);
Ocas;
u obojživelníků :
Jsou přidány krční a sakrální úseky (každý 1 obratel);
u plazů :
Cervikální (8 obratlů, I - atlas, II - osa);
Hrudní (kloubové s žebry, z nichž některá jsou vpředu kloubově spojena s hrudní kostí);
Bederní, sakrální (obsahují větší počet obratlů);
Ocas;
u ptáků :
Cervikální (až 25 obratlů);
Hruď;
Lumbální kost křížová a část ocasních obratlů jsou pevně spojeny a tvoří mohutnou křížovou kost;
u savců :
Cervikální (7 obratlů);
Hrudní (9-24 obratlů);
Bederní (3-9 obratlů);
Sakrální (obratle se spojí);
Kaudální (koccygeální).
U člověka dochází k různým defektům ve formování páteřního kanálu, kterým může spina bifida vystupovat, dále k tvorbě patologických křivek včetně laterálních (skolióza), může být zachována kaudální oblast.
Kostra hlavy se dělí na mozkovou a viscerální lebku. V procesu evoluce došlo ke snížení počtu kostí mozkové lebky, nahrazení chrupavčitých částí kostí a snížení počtu pohyblivých prvků. Postupně se také snižoval počet odbočných oblouků viscerálního skeletu. První a druhý branchiální oblouk se změnily a daly vzniknout čelistem, kostem obličejové lebky, sluchovým kostem a chrupavce hrtanu.
U ryb je oblouk I (čelistní, skládá se z palatokvadratu a Meckelových chrupavek) přímo spojen s obloukem II (skládá se z hyomandibulární chrupavky a hyoidu). Tento typ spojení mezi mozkem a obličejovou lebkou se nazývá hyostyl(protože II oblouk navazuje na hyomandibulární chrupavku). Počínaje obojživelníky se první oblouk se svou palatokvadratní chrupavkou spojuje se základnou lebky. Tento typ připojení se nazývá autostyle. Sloupec sluchové kosti je tvořen z hyomandibulární chrupavky, ze které se následně tvoří třmeny. Při poruchách ontogeneze může mít člověk pouze jednu z těchto sluchových kostí (jako u obojživelníků a plazů) místo tří.
Podívejme se na stavbu sluchového orgánu u různých tříd obratlovců. U Ryba Existuje pouze vnitřní ucho, které zahrnuje membránový labyrint s otolity, polokruhovými kanálky a kochleárním rudimentem.
U obojživelníci Je zde vnitřní ucho stejné stavby a střední ucho s jednou sluchovou kůstkou a bubínkem.
U plazi vnitřní ucho se skládá z hlemýždě a polokruhových kanálků, střední ucho je stejné jako u obojživelníků, je zde rudiment zevního zvukovodu.
U ptactvo zevní zvukovod je dobře formován.
U savců vnitřní ucho zahrnuje hlemýždě 2,5 otáčky, polokruhové kanálky, váček, utrikulum; střední ucho - bubínková dutina, ve které se nacházejí sluchové kůstky - třmen (zbytek hyomandibulární chrupavky), incus (zbytek m. quadratus palatocartilage), malleus (zbytek Meckelovy chrupavky), je bubínek membrána, vnější zahrnuje vnější zvukovod a boltec.
Ve viscerální kostře se tak v procesu evoluce vystřídaly chrupavčité čelisti kostěnými, změnil se typ uchycení čelistí a změnila se funkce prvků útrobních oblouků.
V kostře končetin vznikly pásy s pohyblivě uchycenou kostrou volných končetin; vznikla pětiprstá končetina zemského typu; počet kostí v distálních úsecích se snížil; proximální části končetin byly prodlouženy a distální části zkráceny.
Osoba může mít následující vady: polydaktylie A polyfalanga(zvýšení počtu prstů a falangů); posunutí pásu horních končetin do úrovně I-II hrudních obratlů - Sprengelova nemoc; vrozené anomálie žeber a deformity hrudníku - trychtýř-voda, kýlovitý atd.
FYLOGENEZE SVALŮ
Svaly plní funkci pohybu a dělí se na:
Somatický, který se vyvíjí z myotomů a je inervován míšními nervy (příčně pruhovaný);
Viscerální – vyvíjí se z jiných částí mezodermu, inervován autonomními nervy, může být pruhovaný a hladký.
U dolních strunatců jsou viscerální svaly umístěny v oblasti žaberních oblouků a postupně se z nich formují žvýkací svaly, svaly hltanu, poté svaly šíje a ramenního pletence a také obličejové svaly. . Svaly krku a obličeje mají komplexní inervaci, která je spojena s fázemi fylogeneze. Somatické svaly u dolních strunatců jsou lokalizovány ve formě myomer, dále se tvoří svaly končetin, tvar, velikost a funkce svalů se stávají rozmanitějšími, zvyšuje se jejich inervace, zejména volných končetin u primátů. Svaly trupu jsou v procesu evoluce redukovány.
Úkolystudovat tuto sekci:
- Tato část pomáhá pochopit způsoby evoluce organického světa, postupné komplikace nejen strukturální, ale také fyziologické a biochemické mechanismy, které vedly ke vzniku moderních kraniálních strunatců, včetně primátů a člověka;
- v procesu srovnávání je možné jednoznačně ověřit fyziologickou podmíněnost vzniku nebo komplikace určitých struktur, jakož i jejich vymizení, tj. lépe porozumět stavbě lidského těla;
- pomáhá porozumět lidské embryologii, protože podle základního biogenetického zákona formulovaného v roce 1866 Haeckelem a Müllerem: ontogeneze je krátké a rychlé opakování fylogeneze.
Proto se v embryogenezi lidé vyznačují výskytem struktur, funkcí a biochemických reakcí, které byly přítomny u jejich předchůdců strunatců. Takovým opakováním se říká rekapitulace. V ontogenezi člověka se většinou neobjevují struktury dospělých ryb, obojživelníků a plazů, ale pouze jejich larvy. Navíc rané fáze vývoje rekapitulují úplněji než ty pozdější.
Evoluční vývoj ale není prostým opakováním strukturálních rysů předků, ale předpokládá změny při formování orgánů a systémů . Tento:
- cenogeneze - adaptace embrya na jiné prostředí, které se u dospělých forem ztrácí, např. žábry u pulce; provizorní orgány amniot;
- fylembryogeneze - vývojové změny, které mají adaptivní význam:
A) anabolie (prodloužení) - přidání dalších stupňů, například svalové bránice savců, peří ptáků;
b) odchylka - odchylka od programu ve středních fázích morfogeneze, například tvorba sluchových kůstek, výskyt alveolární struktury plic místo buněčné; biogenetický zákon je v těchto případech částečně naplněn;
PROTI) archalaxa - odchylka ontogeneze hned na začátku, kdy není naplněn biogenetický zákon, např. tvorba ochlupení začíná ztluštěním epidermis, která je ponořena do koria;
G) heterochronie - změna doby tvorby struktury, například časná tvorba placentární amnionu; A heterotopie - změna umístění struktury, například vytvoření lidského ramenního pletence na úrovni krčních obratlů III-IV, který se poté přesune na úroveň I-II hrudních obratlů.
Při poruchách embryogeneze se u dospělých mohou vyvinout příznaky, které byly přítomny u jejich vzdálených předků - atavismy. Pokud snižují životaschopnost, jsou tzv atavistický(nebo rodový z anĉetre) neřesti. Atavismy vznikají kvůli:
Nedostatečný rozvoj orgánů (hypoplazie bránice, rozštěp patra - „rozštěp patra“ atd.);
Perzistence (zachování) embryonálních struktur (patent ductus ductus, laterální krční píštěle, umbilikální píštěle atd.);
Poruchy pohybu orgánů v ontogenezi (pánevní umístění orgánů, kryptorchismus atd.).
Uvažujme umístění strunatců ve světě zvířat a jejich klasifikace.
Kmen Chordata sjednocuje mnohobuněčné deuterostomy různých úrovní organizace. Navíc absolutně všichni strunatci, alespoň v jedné fázi vývoje, mají tři vlastnosti, které je odlišují od ostatních zvířat:
- osový komplex orgánů, který je založen na vnitřní kostře - notochordu, nad ním - nervové trubici, pod ní - trávicí trubici;
- do hltanu prostupují hltanové štěrbiny, které jsou u vyšších strunatců podepřeny útrobními oblouky;
- ocasní část
Kromě toho lze zaznamenat přítomnost žilního systému portální žíly (krev z gastrointestinálního traktu prochází játry) a strukturu pokožky epidermis a dermis, ale vrstva ostnokožců má podobnou strukturu.
Kmen Chordata zahrnuje čtyři podfyla:
I. Hemichordata (Hemichordata) - balianoglossus, ptychodera;
- II. Larvální strunatci (Urochordata) - ascidiánové;
- III. Skullless (Acrania) - lancelety;
- IV. Obratlovci (Vertebrata), které zahrnují několik tříd:
Skupina Anamnie (nižší obratlovci)
- Cyklostomy (Cyclostomata) - mihule, hagfish - nemají čelisti;
- supertřída Ryby:
A. třída Chrupavčité a pancéřové ryby (Chondrichthyes) - žraloci, rejnoci, chiméry;
b. třída Bony fish (Osteichthyes) - ostatní ryby;
- Obojživelníci (Amphibia):
A. Ocasáči - čolci, mloci;
b. Bezocasá zvířata - žáby, ropuchy, rosničky;
PROTI. Beznohé jsou ceciliany;
Skupina Amniota (vyšší obratlovci)
- Plazi (Reptilia):
A. Šupinatý - ještěrky, hadi;
b. Želvy;
PROTI. Krokodýli;
g. Beakheads - hatteria;
- Ptáci (Aves) - vedlejší větev evoluce, mají mnoho specifických adaptací
A. Běžci - pštrosi;
b. tučňáci;
PROTI. Keeleds - ostatní ptáci;
- Savci (Mammalia):
A. Oviparous - ptakopysk, echidna;
b. Vačnatci - klokani;
PROTI. Placenty - ostatní zvířata a lidé.
FYLOGENEZE VNĚJŠÍ INTEGRACE
Hlavní funkce kožní vrstvy jsou vymezení a ochrana mnohobuněčného organismu. Slupka obratlovců získává řadu dalších funkcí. U strunatců se krycí vrstva skládá z:
Epidermis vyvíjející se z ektodermu;
Dermis vyvíjející se z mezodermu.
Směry evoluce jsou následující:
Jednovrstvá epidermis (lanceleta) se stává vícevrstevnou (u obratlovců), vrstvy se diferencují a navenek keratinizují; objevují se deriváty epidermis - zrohovatělé šupiny, drápy, vlasy atd.
Tenká, želatinová pojivová tkáň dermis se zahušťuje a získává na síle v důsledku zvýšení počtu vláken.
Pokožka a dermis mají četné deriváty: chromatofory, šupiny, zrohovatělé štítky, lastury, destičky, zobák, drápy, peří, vlasy, rohy, kopyta. Všechny strunatce mají žlázy. U nižších živočichů jsou to jednobuněčné slizniční žlázy, počínaje rybami se objevují jedovaté žlázy, dále mazové žlázy (u ptáků), u savců se přidávají mléčné a potní žlázy.
Osoba může mít následující poruchy ontogeneze kožního povlaku: albinismus(plné) a vitiligo(neúplná) depigmentace kůže, alopecie(nepřítomnost) a hypertrichóza(nadměrný růst) vlasů, hyperkeratóza A pachyonychie- ztluštění epidermis a nehtů, resp. polythelia A polymastie- zvýšení počtu bradavek a mléčných žláz, resp.
FYLOGENEZE KOSTEL
Nižší strunatci žijí ve vodě, zatímco vyšší strunatci žijí převážně na souši. Přechod z hustšího vodního prostředí do vzdušného vedl k výrazné komplikaci kostry a svalů, zejména kostra končetin a jejich pletenců se stala složitější a zvýšil se počet specializovaných svalů obsluhujících končetiny. Hlavní funkce kostry: podpora, základ pro svalový úpon, účast na minerálním metabolismu.
Na notochordu je založena osová kostra, kolem notochordu se pak vytvořily obratle, ty se diferencovaly, zvětšil se počet částí páteře, u člověka se vlivem vzpřímeného držení těla vytvořily ohyby - 2 kyfózy a 2 lordózy a zbytky v. notochord se zachoval pouze v centrálních úsecích meziobratlových plotének.
Tím pádem, v rybách Existují dvě části páteře:
Trup (trmenové obratle kloubově spojené s žebry);
Ocas;
u obojživelníků :
Jsou přidány krční a sakrální úseky (každý 1 obratel);
u plazů :
Cervikální (8 obratlů, I - atlas, II - osa);
Hrudní (kloubové s žebry, z nichž některá jsou vpředu kloubově spojena s hrudní kostí);
Bederní, sakrální (obsahují větší počet obratlů);
Ocas;
u ptáků :
Cervikální (až 25 obratlů);
Hruď;
Lumbální kost křížová a část ocasních obratlů jsou pevně spojeny a tvoří mohutnou křížovou kost;
u savců :
Cervikální (7 obratlů);
Hrudní (9-24 obratlů);
Bederní (3-9 obratlů);
Sakrální (obratle se spojí);
Kaudální (koccygeální).
U člověka dochází k různým defektům ve formování páteřního kanálu, kterým může spina bifida vystupovat, dále k tvorbě patologických křivek včetně laterálních (skolióza), může být zachována kaudální oblast.
Kostra hlavy se dělí na mozkovou a viscerální lebku. V procesu evoluce došlo ke snížení počtu kostí mozkové lebky, nahrazení chrupavčitých částí kostí a snížení počtu pohyblivých prvků. Postupně se také snižoval počet odbočných oblouků viscerálního skeletu. První a druhý branchiální oblouk se změnily a daly vzniknout čelistem, kostem obličejové lebky, sluchovým kostem a chrupavce hrtanu.
U ryb je oblouk I (čelistní, skládá se z palatokvadratu a Meckelových chrupavek) přímo spojen s obloukem II (skládá se z hyomandibulární chrupavky a hyoidu). Tento typ spojení mezi mozkem a obličejovou lebkou se nazývá hyostyl(protože II oblouk navazuje na hyomandibulární chrupavku). Počínaje obojživelníky se první oblouk se svou palatokvadratní chrupavkou spojuje se základnou lebky. Tento typ připojení se nazývá autostyle. Sloupec sluchové kosti je tvořen z hyomandibulární chrupavky, ze které se následně tvoří třmeny. Při poruchách ontogeneze může mít člověk pouze jednu z těchto sluchových kostí (jako u obojživelníků a plazů) místo tří.
Podívejme se na stavbu sluchového orgánu u různých tříd obratlovců. U Ryba Existuje pouze vnitřní ucho, které zahrnuje membránový labyrint s otolity, polokruhovými kanálky a kochleárním rudimentem.
U obojživelníci Je zde vnitřní ucho stejné stavby a střední ucho s jednou sluchovou kůstkou a bubínkem.
U plazi vnitřní ucho se skládá z hlemýždě a polokruhových kanálků, střední ucho je stejné jako u obojživelníků, je zde rudiment zevního zvukovodu.
U ptactvo zevní zvukovod je dobře formován.
U savců vnitřní ucho zahrnuje hlemýždě 2,5 otáčky, polokruhové kanálky, váček, utrikulum; střední ucho - bubínková dutina, ve které se nacházejí sluchové kůstky - třmen (zbytek hyomandibulární chrupavky), incus (zbytek m. quadratus palatocartilage), malleus (zbytek Meckelovy chrupavky), je bubínek membrána, vnější zahrnuje vnější zvukovod a boltec.
Ve viscerální kostře se tak v procesu evoluce vystřídaly chrupavčité čelisti kostěnými, změnil se typ uchycení čelistí a změnila se funkce prvků útrobních oblouků.
V kostře končetin vznikly pásy s pohyblivě uchycenou kostrou volných končetin; vznikla pětiprstá končetina zemského typu; počet kostí v distálních úsecích se snížil; proximální části končetin byly prodlouženy a distální části zkráceny.
Osoba může mít následující vady: polydaktylie A polyfalanga(zvýšení počtu prstů a falangů); posunutí pásu horních končetin do úrovně I-II hrudních obratlů - Sprengelova nemoc; vrozené anomálie žeber a deformity hrudníku - trychtýř-voda, kýlovitý atd.
FYLOGENEZE SVALŮ
Svaly plní funkci pohybu a dělí se na:
Somatický, který se vyvíjí z myotomů a je inervován míšními nervy (příčně pruhovaný);
Viscerální – vyvíjí se z jiných částí mezodermu, inervován autonomními nervy, může být pruhovaný a hladký.
U dolních strunatců jsou viscerální svaly umístěny v oblasti žaberních oblouků a postupně se z nich formují žvýkací svaly, svaly hltanu, poté svaly šíje a ramenního pletence a také obličejové svaly. . Svaly krku a obličeje mají komplexní inervaci, která je spojena s fázemi fylogeneze. Somatické svaly u dolních strunatců jsou lokalizovány ve formě myomer, dále se tvoří svaly končetin, tvar, velikost a funkce svalů se stávají rozmanitějšími, zvyšuje se jejich inervace, zejména volných končetin u primátů. Svaly trupu jsou v procesu evoluce redukovány.
Fylogeneze orgánových systémů strunatců je posuzována v souladu s progresivním směrem evoluce tohoto druhu živočichů od podkmene lebečních až po třídu savců. Organizace orgánových systémů třídy Ptáci není popsána vzhledem k tomu, že ptáci se vyvinuli z plazů mnohem později než savci a jsou vedlejší větví evoluce strunatců.
Vnější obklady
Závoje Jakékoli zvíře vždy plní funkci vnímání vnějších podráždění a také chrání tělo před škodlivými vlivy prostředí. Intenzifikace první funkce kůže vede v procesu evoluce mnohobuněčných živočichů ke vzniku nervového systému a smyslových orgánů. Intenzifikace druhé funkce je doprovázena diferenciací. Charakteristické je také rozšíření funkcí, v jehož důsledku se kůže jako ochranný orgán podílí i na výměně plynů, termoregulaci a vylučování a výživě potomků. To je způsobeno komplikací struktury kožních vrstev, vzhledu a další transformace četných příloh a žláz.
U všech strunatců má kůže dvojí – ekto- a mezodermální – původ. Z ektodermu se vyvíjí epidermis a z mezodermu dermis. Bezlebečtí se vyznačují slabým stupněm diferenciace obou vrstev kůže. Pokožka je jednovrstevná, válcovitá, obsahuje jednobuněčné slizniční žlázky, škára je volná a obsahuje malý počet buněk pojivové tkáně.
U podtypu Vertebrate se epidermis stává vícevrstevnou, přičemž buňky ve spodní vrstvě se neustále množí a v horních vrstvách se diferencují, odumírají a odlupují se. V dermis se objevují vlákna pojivové tkáně, která dodává kůži pevnost. Kůže tvoří přívěsky, různé v závislosti na životním stylu a úrovni organizace, stejně jako žlázy, které plní různé funkce.
U ryb jsou žlázy v epidermis jednobuněčné. Stejně jako lancelet vylučují hlen, který usnadňuje pohyb ve vodě. Tělo ryb je pokryto šupinami, které mají různé struktury v závislosti na jejich systematické poloze. Šupiny chrupavčitých ryb se nazývají plakoidní. Má tvar hrotu a skládá se z dentinu pokrytého z vnější strany sklovinou (obr. 14.1). Dentin je mezodermálního původu, vzniká fungováním buněk pojivové tkáně, které vyčnívají zvenčí ve formě papily. Sklovina, která je tvrdší nebuněčnou látkou než dentin, je tvořena papilou epidermis a pokrývá vnější stranu plakoidních šupin.
Celý povrch těla chrupavčitých ryb, stejně jako dutina ústní, jejíž sliznice pochází z ektodermu, je pokryta plakoidními šupinami. Přirozeně funkce šupin v dutině ústní souvisí se zachycováním a zadržováním potravy, jsou tedy značně zvětšené a jsou to zuby. Kostnaté ryby mají jiný typ šupin. Vypadá jako tenké kulaté kostní destičky pokryté tenkou vrstvou epidermis. Kostní šupina se vyvíjí zcela na úkor dermis, ale svým původem souvisí s primitivní plakoidní šupinou.
Kůže primitivních vyhynulých obojživelníků - stegocefalů - odpovídala kůži ryb a byla pokryta i šupinami. Moderní obojživelníci mají tenkou, hladkou kůži bez šupin, která se účastní výměny plynů. To je usnadněno přítomností velkého počtu mnohobuněčných slizniční žlázy, jehož tajemství neustále hydratuje pokožku a má baktericidní vlastnosti. Některé kožní žlázy řady obojživelníků se diferencovaly na orgány produkující toxiny, které je chrání před nepřáteli (viz část 23.1).
Rýže. 14.1. Pokládání plakoidních šupin:
1 - buňky tvořící sklovinu, 2- pokožka, 3- smalt, 4- skleroblasty tvořící dentin, 5- dentin, 6- dermální papila
Plazi, kteří zcela přešli do pozemské existence, mají suchou kůži, která se nepodílí na dýchání. Horní vrstva epidermis keratinizuje. Rohové šupiny některých plazů jsou tenké a elastické, zatímco u jiných splývají dohromady a vytvářejí, jako u želv, mocnou rohovitou krunýř. Většina plazů línají, jak rostou, a pravidelně shazují svůj rohový kryt. Moderní plazi nemají kožní žlázy.
Kůže savců je postavena nejsložitěji kvůli rozmanitosti funkcí, které vykonávají. Charakteristické jsou různé deriváty kůže: chlupy, drápy, rohy, kopyta, ale i pot, mazové a mléčné žlázy. Primitivnější savci – hmyzožravci, hlodavci a někteří další – si spolu s chlupy zachovali i zrohovatělé šupiny na ocase. Jejich chlupy rostou v mezerách mezi šupinami, ve skupinách po 3-7. U pokročilejších savců, kteří ztratili šupiny, je zachováno stejné uspořádání srsti (obr. 14.2), pokrývající téměř celé tělo, s výjimkou některých oblastí, jako jsou například chodidla a dlaně člověka.
Vlasyřada savců se rozlišuje na typické, sloužící k termoregulaci, a velké, popř vibrissae, jehož základy jsou spojeny se smyslovými nervovými zakončeními. U většiny savců se vibrissy nacházejí v ústech a nosu, u primátů jsou redukovány v důsledku zvýšené hmatové funkce předních končetin, u mnoha vejcorodých a vačnatců jsou rozptýleny po celém těle. Tato skutečnost může naznačovat, že ochlupení předků savců plnilo především hmatové funkce a poté, jak množství ochlupení přibývalo, začalo se podílet na termoregulaci. Při ontogenezi člověka se tvoří větší množství vlasových pupenů, ke konci embryogeneze však dochází k redukci většiny z nich.
Potní žlázy savci jsou homologní s kožními žlázami obojživelníků. Jejich sekret může být slizovitý a obsahovat bílkoviny a tuk. Některé potní žlázy se u raných savců diferencovaly na mléčné žlázy. U vejcorodých zvířat (platypus, echidna) jsou mléčné žlázy svou stavbou a vývojem podobné žlázám potním. Podél okrajů vyvíjející se bradavky mléčné žlázy lze nalézt postupné přechody z typických potních žláz do mléčných žláz (obr. 14.3). Počet mléčných žláz a bradavek koreluje s plodností (od 25 do jednoho páru), ale v embryogenezi všech savců se na povrchu břicha tvoří „mléčné linie“, táhnoucí se od podpaží až po třísla. Následně se podél těchto linií diferencují bradavky, z nichž většina pak podstoupí redukci a zmizí. V lidské embryogenezi se tedy zpočátku vytvoří pět párů bradavek a následně zůstane pouze jeden (obr. 14.4).

Rýže. 14.4. Embryogeneze přední břišní stěny člověka. A - embryo ve věku 5 týdnů (viditelné mléčné linie); B - diferenciace pěti párů bradavek; V - embryo ve věku 7 týdnů

Rýže. 14.5. Atavistické anomálie vývoje kůže.
A - hypertrichóza; B - polymastie
Mazové žlázy se tvoří v kůži pouze u savců. Jejich sekrece, promašťující vlasy a povrch pokožky, jim dodává nesmáčení a pružnost.
Ontogeneze kůže a úponů kůže savců a lidí odráží jejich vývoj podle typu archalaxis. Ve skutečnosti ani základy rohovitých šupin charakteristické pro plazy, ani dřívější formy kožních přívěsků nerekapitulují v jejich embryogenezi. Současně se ve fázi sekundární organogeneze okamžitě vyvinou základy vlasových folikulů. Poruchy rané ontogeneze lidské kůže mohou způsobit výskyt některých drobných atavistických malformací: hypertrichóza (zvýšený růst ochlupení), polytelie (zvýšený počet bradavek), polymastie (zvýšený počet mléčných žláz) (obr. 14.5). Všechny jsou spojeny s porušením redukce nadměrného počtu těchto struktur a odrážejí evoluční spojení člověka s nejbližšími formami předků - savci. To je důvod, proč je nemožné, aby se lidem a jiným savcům narodilo potomstvo s atavistickými rysy kůže charakteristickými pro vzdálenější předky. Jedním z nejznámějších příznaků nedonošenosti u novorozenců je zvýšený růst kožního ochlupení. Krátce po narození přebytečné vlasy obvykle vypadnou a vlasové folikuly se zmenšují.
SVALOVÝ SYSTÉM
Fylogeneze motorických funkcí je základem progresivního vývoje zvířat. Úroveň jejich organizace proto primárně závisí na povaze motorické činnosti, která je určena charakteristikami organizace muskuloskeletální systém, prodělaly velké evoluční přeměny ve kmeni Chordata v důsledku změn stanovišť a změn forem pohybu. Ve vodním prostředí živočichů, kteří nemají exoskeleton, totiž dochází k monotónním pohybům v důsledku ohýbání celého těla, zatímco život na souši je pro jejich pohyb pomocí končetin příznivější.
Podívejme se samostatně na vývoj kostry a svalového systému.
Kostra
V strunatcích vnitřní kostra. Podle stavby a funkcí se dělí na osovou kostru, kostru končetin a hlavu.
Axiální skelet
V podtypu Skullless existuje pouze axiální skelet ve formě akordu. Je postaven z vysoce vakuolizovaných buněk, těsně přiléhajících k sobě a pokrytých na vnější straně společnými elastickými a vazivovými membránami. Pružnost tětivy je dána turgorovým tlakem jejích buněk a pevností membrán. Notochord se tvoří v ontogenezi všech strunatců a u více organizovaných zvířat plní ne tak podpůrnou, ale morfogenetickou funkci, je orgánem, který provádí embryonální indukci.
Po celý život u obratlovců se notochord zachovává pouze v cyklostomech a některých nižších rybách. U všech ostatních zvířat je snížena. U lidí jsou v postembryonálním období zachovány základy chordy ve formě meziobratlových plotének nucleus pulposus. Zachování nadměrného množství notochordálního materiálu, když je narušena jeho redukce, je zatíženo možností vzniku nádorů u lidí - akord, vznikající na jejím základě.
U všech obratlovců je notochord postupně nahrazován obratel, vyvíjející se ze sklerotomů somitů a je funkčně nahrazen páteř. Toto je jeden z nejjasnějších příkladů homotopické orgánové substituce (viz § 13.4). Tvorba obratlů ve fylogenezi začíná vývojem jejich oblouků, které pokrývají neurální trubici a stávají se místy svalového připojení. Počínaje chrupavčitými rybami se zjišťuje chrupavčitost schránky notochordu a růst základů obratlových oblouků, v důsledku čehož se tvoří obratlová těla. Srůstem horních obratlových oblouků nad neurální trubicí vznikají trnové výběžky a páteřní kanál, který uzavírá neurální trubici (obr. 14.6).

Rýže. 14.6. Vývoj obratlů. A-rané stadium; B- další fáze:
1 -akord, 2- akordová skořápka, 3- horní a dolní oblouky obratlů, 4- trnový výběžek, 5- osifikační zóny, 6-notochordový rudiment, 7 - chrupavčité tělo obratle
Nahrazení notochordu páteří - výkonnějším podpůrným orgánem se segmentovou strukturou - umožňuje zvětšit celkovou velikost těla a aktivuje motorické funkce. Další progresivní změny v páteři jsou spojeny s tkáňovou substitucí - náhradou chrupavčité tkáně kostí, která se nachází u kostnatých ryb, a také s její diferenciací na úseky.
Ryby mají pouze dvě části páteře: kmen A ocas. To je způsobeno jejich pohybem ve vodě v důsledku ohýbání těla.
Obojživelníci také získávají krční A křížový oddělení, z nichž každé představuje jeden obratel. První poskytuje větší pohyblivost hlavy a druhý poskytuje podporu zadním končetinám.
U plazů se prodlužuje krční páteř, jejíž první dva obratle jsou pohyblivě spojeny s lebkou a zajišťují větší pohyblivost hlavy. Objeví se bederníúsek ještě slabě ohraničený od hrudního a křížová kost se skládá již ze dvou obratlů.
Savci se vyznačují stabilním počtem obratlů v krční oblasti, rovným 7. Pro velký význam v pohybu zadních končetin je křížová kost tvořena 5-10 obratli. Bederní a hrudní oblast jsou od sebe jasně ohraničeny.
U ryb nesou všechny obratle trupu žebra, která nejsou srostlá navzájem ani s hrudní kostí. Dodávají tělu stabilní tvar a poskytují oporu svalům, které ohýbají tělo v horizontální rovině. Tato funkce žeber je zachována u všech obratlovců, kteří provádějí hadí pohyby – u ocasatých obojživelníků a plazů se proto jejich žebra nacházejí také na všech obratlech kromě ocasních.
U plazů část hrudních žeber splývá s hrudní kostí a tvoří hrudník a u savců hrudník zahrnuje 12-13 párů žeber.

Rýže. 14.7. Vývojové anomálie osového skeletu. A - zbytková krční žebra (znázorněna šipkami); B - Nesrůstání trnových výběžků obratlů v hrudní a bederní oblasti. Spina bifida
Ontogeneze osového skeletu člověka rekapituluje hlavní fylogenetická stádia jeho vzniku: v období neurulace vzniká notochord, který je následně nahrazen chrupavčitým a následně kostěným páteřem. Na krčních, hrudních a bederních obratlích se vyvíjí pár žeber, načež se redukují krční a bederní žebra a hrudní žebra splývají vpředu mezi sebou a s hrudní kostí a tvoří hrudní koš.
Narušení ontogeneze axiálního skeletu u člověka se může projevit takovými atavistickými vývojovými vadami, jako je nesrůstání trnových výběžků obratlů, což má za následek vznik spinabifida - defekt páteřního kanálu. V tomto případě mozkové pleny často pročnívají defektem a a rozštěp páteře(obr. 14.7).
Ve věku 1,5-3 měsíce. Lidské embryo má kaudální páteř sestávající z 8-11 obratlů. Porušení jejich redukce následně vysvětluje možnost výskytu tak známé anomálie osového skeletu jako je vytrvalost ocasu.
Porušení redukce krčních a bederních žeber je základem jejich zachování v postnatální ontogenezi.
Kostra hlavy
Přední pokračování axiálního skeletu je axiální, nebo mozek, lebka, slouží k ochraně mozku a smyslových orgánů. Rozvíjí se vedle ní viscerální, nebo obličejová lebka, tvoří oporu pro přední část trávicí trubice. Obě části lebky se vyvíjejí odlišně a z různých rudimentů. V raných fázích evoluce a ontogeneze spolu nejsou spojeny, ale později toto spojení vzniká.

Rýže. 14.8. Lidská lebka s metodickým stehem (označeno šipkou)
V zadní části axiální lebky během vývoje jsou nalezeny stopy segmentace, proto se má za to, že je výsledkem fúze anlage předních obratlů mezi sebou. Složení mozkové lebky také zahrnuje analáž chrupavčitých pouzder mezenchymálního původu obklopujících orgány sluchu, čichu a zraku. Navíc část mozkové lebky (ležící před sella turcica), která nemá segmentaci, se zjevně vyvíjí jako novotvar v důsledku zvětšení velikosti předního mozku.
Fylogeneticky prošla mozková lebka třemi fázemi vývoje: membranózní, chrupavčitý A kost.
V cyklostomech je téměř celá membránová a nemá přední, nesegmentovanou část.
Lebka chrupavčitých ryb je téměř celá chrupavčitá a zahrnuje jak zadní, primárně segmentovanou část, tak přední část.
U kostnatých ryb a jiných obratlovců se axiální lebka stává kostnatou v důsledku procesů osifikace chrupavky v oblasti její základny (bazální, sfenoidní, etmoidní kosti) a v důsledku vzhledu krycích kostí v její horní části (temenní , čelní, nosní kosti). Kosti axiální lebky procházejí oligomerizací v procesu progresivní evoluce. Tomu nasvědčuje výskyt velkého počtu osifikačních zón a jejich následné splynutí během tvorby takových kostí, jako je frontální, temporální atd. Široce známé u lidí jsou takové anomálie mozkové lebky jako přítomnost interparietálních kostí a také dvou čelních kostí s metopickým stehem mezi nimi (obr. 14.8). Nejsou provázeny žádnými patologickými jevy, a proto jsou většinou náhodně objeveny až po smrti.
U nižších obratlovců se také poprvé objevuje viscerální lebka. Vzniká z mezenchymu ektodermálního původu, který je seskupen ve formě obloukovitých kondenzátů v prostorech mezi žaberními štěrbinami hltanu. První dva oblouky dostávají zvláště silný vývoj a dávají vzniknout čelistním a jazylkovým obloukům dospělých zvířat. Následující oblouky, čítající 4-5 párů, plní podpůrnou funkci pro žábry a jsou tzv žábry.
U chrupavčitých ryb jsou před čelistním obloukem obvykle ještě 1-2 páry premaxilárních oblouků, které jsou rudimentární povahy. To ukazuje, že předci obratlovců měli větší počet viscerálních oblouků než 6 nebo 7 a k jejich diferenciaci došlo na pozadí oligomerizace.
Čelistní oblouk se skládá ze dvou chrupavek. Horní se nazývá palatoquadrate, on plní funkci primární horní čelisti. Nižší, popř Meckel, chrupavka - primární dolní čelist. Na ventrální straně hltanu jsou Meckelovy chrupavky navzájem spojeny tak, že čelistní oblouk uzavírá dutinu ústní do prstence. Druhý viscerální oblouk na každé straně tvoří hyomandibulární chrupavka srostlá se spodinou lebeční a jazylka spojená s Meckelovou chrupavkou. U chrupavčitých ryb jsou tedy obě primární čelisti spojeny s axiální lebkou prostřednictvím druhého viscerálního oblouku, ve kterém hyomandibulární chrupavka působí jako závěs na mozkovou lebku. Tento typ spojení mezi čelistmi a axiální lebkou se nazývá hyostyl(obr. 14.9).
U kostnatých ryb začíná výměna primárních čelistí za sekundární, skládající se z falešných kostí - čelist a premaxilla nahoře a zub dole. Palatoquadrát a Meckelovy chrupavky se zmenšují a pohybují se dozadu. Hyomandibulární chrupavka nadále funguje jako závěs, takže lebka zůstává hyostylní.
Obojživelníci v souvislosti s přechodem k pozemské existenci prodělali výrazné změny na viscerální lebce. Větevné oblouky jsou částečně redukovány a částečně se změnou své funkce stávají součástí chrupavčitého aparátu hrtanu. Čelistní oblouk se svým horním prvkem - palatinovou kvadrátní chrupavkou - zcela splyne se spodinou mozkové lebky a lebka se tak stává autostyle. Hyomandibulární chrupavka, značně zmenšená a zbavená funkce zavěšení, umístěná v oblasti první žaberní štěrbiny uvnitř sluchového pouzdra, převzala funkci sluchové kůstky - sloupku - přenášejícího zvukové vibrace z vnějšího do vnitřního ucha. .
Viscerální lebka plazů je také autostyle. Čelisťový aparát se vyznačuje vyšším stupněm osifikace než u obojživelníků. Část chrupavčitého materiálu žaberních oblouků je součástí nejen hrtanu, ale i průdušnice.
Dolní čelist savců se kloubí se spánkovou kostí složitým kloubem, který umožňuje nejen zachycení potravy, ale i složité žvýkací pohyby.
Jedna sluchová kůstka - sloupec,- charakteristické pro obojživelníky a plazy, zmenšující se velikost, přechází v stapes, a základy palatokvadrátu a Meckelových chrupavek, které zcela opouštějí čelistní aparát, se přeměňují na kovadlina A kladivo. Vzniká tak jeden funkční řetězec tří sluchových kůstek ve středním uchu, charakteristický pouze pro savce (obr. 14.9).

Rýže. 14.9. Evoluce prvních dvou viscerálních větevních oblouků obratlovců.
A- chrupavčité ryby; B- obojživelník; V- plaz; G- savec:
1 -palatokvadrátová chrupavka, 2-Meckelova chrupavka, 3- hyomandibulární chrupavka, 4-hyoidní, 5- sloupec, 6- překryvné kosti sekundárních čelistí, 7-kovadlina, 8- stapes, 9- kladivo; homologní útvary jsou označeny odpovídajícím stínováním
K rekapitulaci hlavních fází fylogeneze viscerální lebky dochází i v lidské ontogenezi. Porušení diferenciace elementů maxilárního žaberního oblouku do sluchových kůstek je mechanismem pro vznik takové malformace středního ucha, jako je lokalizace v bubínkové dutině pouze jedné sluchové kůstky - sloupku, který odpovídá struktura zařízení pro přenos zvuku obojživelníků a plazů.
Kostra končetiny
Strunatci mají nepárové a párové končetiny. Nepárové (hřbetní, ocasní a řitní ploutve) jsou hlavními orgány pohybu obojživelníků, ryb a v menší míře i obojživelníků ocasatých.Rybám se také vyvíjejí párové končetiny - prsní a pánevní ploutve, na jejichž základě se párové končetiny suchozemských následně se vyvinou čtyřnožci.
Podívejme se blíže na vznik a vývoj párových končetin.
U rybích larev, stejně jako u moderních bezlebkových ryb, postranní kožní záhyby tzv metapleurální(obr. 14.10). Nemají ani kostru, ani vlastní svaly, plní pasivní roli - stabilizují polohu těla a zvětšují plochu břišní plochy, usnadňují pohyb ve vodním prostředí. Pravděpodobně u předků ryb přecházejících k aktivnějšímu životnímu stylu se v těchto záhybech objevily svalové prvky a chrupavčité paprsky, původem spojené se somity a tedy lokalizované metamericky. Takové záhyby, které získaly pohyblivost, mohou působit jako hloubková kormidla, avšak pro změnu polohy těla v prostoru mají větší význam jejich přední a zadní sekce, protože jsou nejvzdálenější od těžiště. Proto evoluce šla cestou zintenzivnění funkcí vnějších částí a oslabení funkcí centrálních částí.

Rýže. 14.10. Tvorba předních a zadních končetin z metapleurálních záhybů: já-III-hypotetická stádia evoluce
V důsledku toho se z předních částí záhybů vyvinuly prsní ploutve a ze zadních ploutve břišní (obr. 14.10). Je možné, že vytvoření pouze dvou párů končetin na bočních stranách těla předcházel rozpad souvislých záhybů do řady párových ploutví, z nichž byla důležitější i přední a zadní. Svědčí o tom existence fosilních pozůstatků nejstarších málo organizovaných ryb s četnými ploutvemi (obr. 14.11). V důsledku splynutí základů chrupavčitých paprsků, brachiální A pánevního pletence. Odpočinek jejich oblasti se diferencovaly na kostra volných končetin.

Rýže. 14.11. Starobylá ryba podobná žralokům s četnými párovými končetinami
U většiny ryb je kostra párových ploutví rozdělena na proximální část, sestávající z malého počtu chrupavčitých nebo kostěných plátů, a na distální část, která zahrnuje velké množství radiálně segmentovaných paprsků. Ploutve jsou spojeny s pletenci končetin nečinně. Nemohou sloužit jako opora pro tělo při pohybu po dně nebo souši. U lalokoploutvých ryb má kostra párových končetin odlišnou stavbu. Celkový počet jejich kostních prvků je snížen a jsou větší. Proximální úsek se skládá pouze z jednoho velkého kostního elementu, který odpovídá humeru nebo femuru předních nebo zadních končetin. Následují dvě menší kosti, homologní s loketní kostí a radiem nebo holenní a lýtkovou kostí. Na nich spočívá 7-12 radiálně umístěných paprsků. Ve spojení s pletenci končetin takové ploutve se jedná pouze o homology pažní nebo stehenní, proto jsou ploutve lalokoploutvých aktivně pohyblivé (obr. 14.12, A, B) a lze jej použít nejen ke změně směru pohybu ve vodě, ale také k pohybu po pevném substrátu.
Život těchto ryb v mělkých vysychajících nádržích v období devonu přispěl k selekci forem s vyvinutějšími a pohyblivějšími končetinami. Přítomnost dalších dýchacích orgánů v nich (viz část 14.3.4) se stala druhým předpokladem pro dosažení pevniny a vzniku dalších adaptací na suchozemskou existenci, což mělo za následek vznik obojživelníků a celé skupiny Tetrapoda. Jejich první zástupci - stegocefálové - měli sedmiprsté a pětiprsté končetiny, které si zachovaly podobnost s ploutvemi lalokoploutvých ryb (obr. 14.12, Obr. B)

Rýže. 14.12. Kostra končetiny lalokoploutvé ryby ( A), jeho základna ( B) a kostra přední tlapy Stegocephala ( V):já- humerus, 2-ulna, 3- poloměr
V kostře zápěstí je zachováno správné radiální uspořádání kostních elementů ve 3-4 řadách, v metakarpu je 7-5 kostí a pak také falangy 7-5 prstů leží radiálně.
U moderních obojživelníků je počet prstů na končetinách pět nebo jsou oligomerizovány na čtyři.
Další progresivní přeměna končetin se projevuje zvýšením stupně pohyblivosti kostních kloubů, snížením počtu kostí v zápěstí, nejprve na tři řady u obojživelníků a poté na dvě u plazů a savců. Současně se také snižuje počet falangů prstů. Charakteristické je také prodloužení proximálních částí končetiny a zkrácení distálních částí.
V průběhu evoluce se mění i uspořádání končetin. Pokud jsou u ryb prsní ploutve na úrovni prvního obratle a jsou vytočené do stran, pak u suchozemských obratlovců v důsledku komplikace orientace v prostoru vzniká krk a pohyblivost hlavy a u plazů a zejména savci se díky vyvýšení těla nad zem pohybují přední končetiny dozadu a jsou orientovány nikoli horizontálně, ale vertikálně. Totéž platí pro zadní končetiny.
Různorodost životních podmínek, které poskytuje suchozemský způsob života, poskytuje rozmanité formy pohybu: skákání, běh, plazení, létání, kopání, lezení po skalách a stromech a při návratu do vodního prostředí plavání. U suchozemských obratlovců lze proto nalézt jak téměř neomezenou rozmanitost končetin, tak jejich úplnou sekundární redukci a opakovaně konvergentně vznikalo mnoho podobných končetinových adaptací v různých prostředích (obr. 14.13). Během procesu ontogeneze však většina suchozemských obratlovců vykazuje společné rysy ve vývoji končetin: tvorba jejich primordií ve formě špatně diferencovaných záhybů, tvorba šesti nebo sedmimístných primordií na ruce a noze, nejvzdálenější které jsou brzy redukovány a následně se vyvine pouze pět (obr. 14.14).

Rýže. 14.13. Kostra přední končetiny suchozemských obratlovců. A-žába- B-mlok; V-krokodýl; G-netopýr; D-Člověk: 1 - humerus, 2-radiusová kost, 3 - zápěstní kosti, 4 - metakarpus, 5 - falangy prstů, 6 - loketní kost
Rýže. 14.14. Stavba vyvíjející se končetiny obratlovce: pp - prepollex, pin - postminimus - další rudimentární prsty I a VII
Je zajímavé, že v embryogenezi vyšších obratlovců rekapituluje nejen stavba končetin předků, ale i proces jejich heterotopie. U lidí se tedy horní končetiny tvoří na úrovni 3.-4. krčního obratle a dolní - na úrovni bederních obratlů. Současně dostávají končetiny inervaci z odpovídajících částí míchy. Heterotopie končetin je doprovázena tvorbou krčních, bederních a sakrálních nervových pletení, jejichž nervy jsou spojeny jednak s těmi segmenty míchy, ze kterých vyrostly v době vzniku končetin a na druhé straně končetin, které se přesunuly na nové místo (obr. 14.15; viz také část 14.2.2.2).
V ontogenezi člověka jsou možné četné poruchy vedoucí ke vzniku atavistických vrozených vývojových vad končetin. Tak, polydaktylie, nebo zvýšení počtu prstů, zděděné jako autozomálně dominantní rys, je výsledkem vývoje analáží dalších prstů, které jsou normálně charakteristické pro vzdálené formy předků. Fenomén polyfalangy je známý, charakterizovaný zvýšením počtu falangů, obvykle palce. Jeho výskyt je založen na vývoji tří článků v prvním prstu, jak je běžně pozorováno u plazů a obojživelníků s nediferencovanými prsty. Bilaterální polyfalanga se dědí autozomálně dominantním způsobem.
Závažnou malformací je porušení heterotopie pletence horní končetiny od krční oblasti až po úroveň 1.-2. hrudního obratle. Tato anomálie se nazývá Sprengelova nemoc nebo vrozené vysoké postavení lopatky (obr. 14.16). Vyjadřuje se tím, že ramenní pletenec na jedné nebo obou stranách je o několik centimetrů vyšší než normální poloha. Vzhledem k tomu, že taková porucha je často doprovázena anomáliemi žeber, hrudní páteře a deformací lopatek, je třeba myslet na to, že mechanismy jejího vzniku nejsou pouze porušením pohybu orgánů, ale také výsledným porušení morfogenetických korelací (viz § 13.4).
Srovnávací anatomický přehled vývoje kostry strunatců ukazuje, že lidská kostra je zcela homologní s podpůrným aparátem rodových a příbuzných forem. Proto lze mnohé malformace jeho vývoje u lidí vysvětlit příbuzností savců s plazy, obojživelníky a rybami. V procesu antropogeneze se však objevily kosterní znaky, které jsou charakteristické pouze pro člověka a jsou spojeny s jeho vzpřímeným držením těla a pracovní aktivitou. Patří mezi ně: 1) změny na chodidle, které přestalo plnit úchopovou funkci, vyjádřené ztrátou schopnosti oponovat palec na noze a vzhled jeho oblouků, které slouží k tlumení nárazů při chůzi; 2) změny páteře - její ohyb ve tvaru S, poskytující plasticitu pohybů ve vertikální poloze; 3) změny v lebce - prudké snížení její obličejové části a zvýšení mozku, přední posunutí foramen magnum, zvýšení mastoidního procesu a vyhlazení týlního reliéfu, ke kterému jsou šíjové svaly a šíjové vazy připojený; 4) specializace horních (předních) končetin jako orgánu porodního; 5) výskyt bradového výběžku v souvislosti s rozvojem artikulované řeči.

Rýže. 14.15. Vznik hrudních končetin, jejich heterotopie a inervace v ontogenezi člověka. A-vrůstání cervikálních myotomů do vyvíjející se přední končetiny embrya; B-vývoj kožní inervace ruky; V- umístění cervikálních a brachiálních plexů zapojených do inervace paže:
1 - cervikální myotomy, 2- hrudní myotomy, 3 - bederní myotomy; písmena C, T, L označují krční, hrudní a bederní segmenty

Rýže. 14.16. Sprengelova nemoc (vysvětlení viz text)
Navzdory tomu, že formování anatomických a morfologických znaků lidské kostry je zřejmě dokončeno, jsou adaptace na vzpřímenou chůzi u člověka, jako všechny adaptace obecně, relativní povahy. Při těžké fyzické aktivitě je tedy možné posunutí obratlů nebo meziobratlových plotének. Po přechodu na vzpřímenou chůzi člověk ztratil schopnost rychle běžet a pohybuje se mnohem pomaleji než většina čtyřnohých zvířat.
Přirozeně, během nitroděložního vývoje se kosterní rysy, které charakterizují člověka jako jedinečný biologický druh, formují v jeho konečných fázích nebo dokonce, jako je páteř ve tvaru S, v časném postnatálním období vývoje. Jsou to vlastně anabolika, která vznikla během fylogeneze primátů. Proto atavistické anomálie kostra spojená se zpožděním ve vývoji vlastností charakteristických pouze pro člověka jsou nejčastější. Prakticky nesnižují vitalitu, ale děti s nimi potřebují ortopedickou korekci, gymnastiku a masáže. Mezi takové anomálie patří mírné formy vrozených plochých nohou, paličkovité nohy, úzký hrudník, absence bradového výběžku a některé další.
Svalová soustava
U zástupců kmene Chordata se svaly dělí podle charakteru vývoje a inervace na somatické a viscerální.
Somatické svalstvo se vyvíjí z myotomů a je inervován nervy, jejichž vlákna vystupují z míchy jako součást ventrálních kořenů míšních nervů. Viscerální svaly se vyvíjí z jiných částí mezodermu a je inervován nervy autonomního nervového systému. Všechny somatické svaly jsou příčně pruhované a viscerální svaly mohou být příčně pruhované nebo hladké (obr. 14.17).

Rýže. 14.17. Somatické a viscerální svaly obratlovců:
1 - somatické svaly vyvíjející se z myotomů, 2- viscerální svaly oblasti žáber
Viscerální svaly
Nejvýraznější změny doznaly viscerální svaly spojené s viscerálními oblouky přední části trávicí trubice. U nižších obratlovců většinu tohoto svalu zastupuje obecný konstriktor viscerálního aparátu - m. constrictor superficialis, pokrývající celou plochu žaberních oblouků na všech stranách. V oblasti čelistního oblouku je tento sval inervován trojklaného nervu(V), v oblasti hyoidního oblouku - obličeje(VII), v oblasti prvního žaberního oblouku - glosofaryngeální(IX), konečně jeho část ležící více kaudálně - putování nerv (X). V tomto ohledu jsou všechny deriváty odpovídajících viscerálních oblouků a s nimi spojené svaly následně inervovány u všech obratlovců uvedenými nervy.
V přední části kompresoru se nachází velká svalová hmota, která slouží čelistnímu aparátu. Za viscerálním aparátem se diferencuje m. trapezius. trapezius, připojené v samostatných svazcích k poslední žaberní štěrbině a přední hraně hřbetní části pletence ramenního. Část povrchového zúžení v oblasti hyoidního oblouku u plazů roste, pokrývá krk zespodu a ze stran a tvoří cervikální zúžení m. sphincter colli. U savců je tento sval rozdělen na dvě vrstvy: hlubokou a povrchovou. Hluboký si zachovává stejný název a povrchový se nazývá platysma myoides a nachází se subkutánně. Tyto dva svaly rostou po celé oblasti hlavy a dávají vzniknout složitému systému obličejových podkožních svalů, který se u primátů a lidí nazývá tzv. napodobit. Proto všechny napodobují
PŘEDNÁŠKA MYOLOGIE FYLOGENEZE, ONTOGENEZE A FUNKČNÍ ANATOMIE SVALOVÉHO SYSTÉMU Účinkuje: Vladimirova Ya. B. Kokoreva T. V.

 Svaly neboli svaly (z latinského musculus - myš, malá myš) jsou orgány těla zvířat a lidí, skládající se z elastické, elastické svalové tkáně, schopné kontrakcí pod vlivem nervových vzruchů. Navrženo k provádění různých akcí: pohyby těla, kontrakce hlasivek, dýchání. Svaly se skládají z 86,3 % vody. V lidském těle je 640 svalů
Svaly neboli svaly (z latinského musculus - myš, malá myš) jsou orgány těla zvířat a lidí, skládající se z elastické, elastické svalové tkáně, schopné kontrakcí pod vlivem nervových vzruchů. Navrženo k provádění různých akcí: pohyby těla, kontrakce hlasivek, dýchání. Svaly se skládají z 86,3 % vody. V lidském těle je 640 svalů
 Motivace: - - - možnosti prováděného pohybu, objem pohybu; aktivní nebo pasivní pohyby jsou spouštěny jednou nebo jinou svalovou skupinou; ovlivněním svalového systému měníme celkový stav; svalový reliéf je vodítkem pro topografii krevních cév a nervů; svalovou transplantaci, to znamená, že sval lze „přeučit“.
Motivace: - - - možnosti prováděného pohybu, objem pohybu; aktivní nebo pasivní pohyby jsou spouštěny jednou nebo jinou svalovou skupinou; ovlivněním svalového systému měníme celkový stav; svalový reliéf je vodítkem pro topografii krevních cév a nervů; svalovou transplantaci, to znamená, že sval lze „přeučit“.
 Vývoj svalů kraniálního původu - z hlavových myotomů (sklerotomů) a mezenchymu větvích oblouků. Inervovány větvemi hlavových nervů míšního původu - z myotomů kmene embrya: z ventrálních myotomů jsou inervovány předními větvemi SMN; - z dorzálních myotomů jsou inervovány zadními větvemi SMN - Autochtonní svaly - svaly, které zůstávají v místě jejich primární anlage. Truncofugální svaly jsou svaly, které se přesunuly z trupu na končetiny. Trunkopetální svaly jsou svaly, které se přesunuly z končetin na trup.
Vývoj svalů kraniálního původu - z hlavových myotomů (sklerotomů) a mezenchymu větvích oblouků. Inervovány větvemi hlavových nervů míšního původu - z myotomů kmene embrya: z ventrálních myotomů jsou inervovány předními větvemi SMN; - z dorzálních myotomů jsou inervovány zadními větvemi SMN - Autochtonní svaly - svaly, které zůstávají v místě jejich primární anlage. Truncofugální svaly jsou svaly, které se přesunuly z trupu na končetiny. Trunkopetální svaly jsou svaly, které se přesunuly z končetin na trup.



 Pruhované hladké 1. Organizační jednotkou je myocyt. Délka asi 50 mikronů. Šířka od 6 mikronů. 2. Mimovolná kontrakce Řízení autonomním nervovým systémem Pohyb je vlnový, pracuje pomalu, jelikož se nervové vlákno NEpřibližuje ke každé buňce Vstupují do činnosti pomalu, ale přetrvávají dlouhou dobu Nemá přesnou prostorovou orientaci buněk 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Srdeční Organizační jednotkou je svalové vlákno - soubor myoblastů plovoucích ve společné cytoplazmě (sarkoplazmě). Mají společné sarkolema. Délka cca 40 -100 mm. Šířka od 7 mm. Dobrovolná kontrakce Ovládání somatickým nervovým systémem Rychlá kontrakce, rychlá reakce, takže každé svalové vlákno má neuromuskulární synapsi Zapnout rychle, ale mít krátkodobý účinek Jasná orientace svalových vláken
Pruhované hladké 1. Organizační jednotkou je myocyt. Délka asi 50 mikronů. Šířka od 6 mikronů. 2. Mimovolná kontrakce Řízení autonomním nervovým systémem Pohyb je vlnový, pracuje pomalu, jelikož se nervové vlákno NEpřibližuje ke každé buňce Vstupují do činnosti pomalu, ale přetrvávají dlouhou dobu Nemá přesnou prostorovou orientaci buněk 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Srdeční Organizační jednotkou je svalové vlákno - soubor myoblastů plovoucích ve společné cytoplazmě (sarkoplazmě). Mají společné sarkolema. Délka cca 40 -100 mm. Šířka od 7 mm. Dobrovolná kontrakce Ovládání somatickým nervovým systémem Rychlá kontrakce, rychlá reakce, takže každé svalové vlákno má neuromuskulární synapsi Zapnout rychle, ale mít krátkodobý účinek Jasná orientace svalových vláken

 Mezi svalovými vlákny jsou tenké vrstvy volného vazivového vaziva – endomysium. Jsou do ní vetkána kolagenová vlákna vnější vrstvy bazální membrány, což napomáhá ke spojení sil při kontrakci myosymplastů. Silnější vrstvy volné pojivové tkáně obklopují několik svalových vláken, tvoří perimysium a rozdělují sval do snopců. Několik svazků je spojeno do větších skupin, oddělených silnějšími vrstvami pojivové tkáně. Pojivová tkáň obklopující povrch svalu se nazývá epimysium.
Mezi svalovými vlákny jsou tenké vrstvy volného vazivového vaziva – endomysium. Jsou do ní vetkána kolagenová vlákna vnější vrstvy bazální membrány, což napomáhá ke spojení sil při kontrakci myosymplastů. Silnější vrstvy volné pojivové tkáně obklopují několik svalových vláken, tvoří perimysium a rozdělují sval do snopců. Několik svazků je spojeno do větších skupin, oddělených silnějšími vrstvami pojivové tkáně. Pojivová tkáň obklopující povrch svalu se nazývá epimysium.
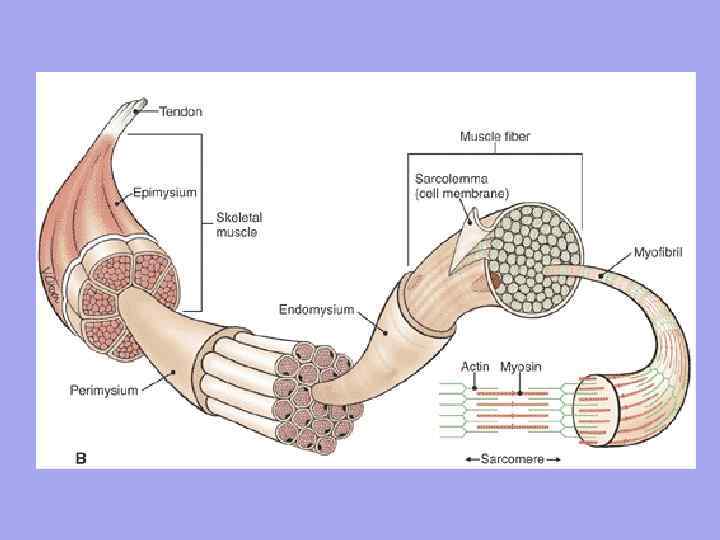
 Sval jako orgán obsahuje pojivovou tkáň Endomysium je tenká pojivová tkáň, která obklopuje každé svalové vlákno a malé skupiny vláken. Perimysium – pokrývá větší komplexy svalových vláken a svalových snopců.
Sval jako orgán obsahuje pojivovou tkáň Endomysium je tenká pojivová tkáň, která obklopuje každé svalové vlákno a malé skupiny vláken. Perimysium – pokrývá větší komplexy svalových vláken a svalových snopců.
 Význam endomysia a perimysia 1. Přes endomysium a perimysium se cévy a nervy přibližují ke svalovému vláknu. Tvoří stroma orgánu; 2. Svalová vlákna se formují do snopců, snopce do svalů; 3. Vzhledem k tomu, že endomysium je srostlé se sarkolemou svalového vlákna, může se kontrahující svalové vlákno natáhnout jen do určité hranice
Význam endomysia a perimysia 1. Přes endomysium a perimysium se cévy a nervy přibližují ke svalovému vláknu. Tvoří stroma orgánu; 2. Svalová vlákna se formují do snopců, snopce do svalů; 3. Vzhledem k tomu, že endomysium je srostlé se sarkolemou svalového vlákna, může se kontrahující svalové vlákno natáhnout jen do určité hranice
 Myofibrily ve vláknu jsou obklopeny obalem – sarkolemou, a ponořeny do speciálního média – sarkoplazmy. Podle obsahu pigmentu a kyslíku se vlákna dělí na bílá a červená. Bílá vlákna jsou anaerobní, obsahují více myofibril a méně sarkoplazmy. Rychle se spouštějí, ale nemohou dlouho fungovat. Příklad: sternocleidomastoideus, m. gastrocnemius. Červená vlákna jsou silná vlákna. V sarkoplazmě je hodně myoglobinu a v mitochondriích cytochromu, ale méně myofibril. Pomalý začátek, ale vydrží dlouho. Příklad: zádové svaly, bránice.
Myofibrily ve vláknu jsou obklopeny obalem – sarkolemou, a ponořeny do speciálního média – sarkoplazmy. Podle obsahu pigmentu a kyslíku se vlákna dělí na bílá a červená. Bílá vlákna jsou anaerobní, obsahují více myofibril a méně sarkoplazmy. Rychle se spouštějí, ale nemohou dlouho fungovat. Příklad: sternocleidomastoideus, m. gastrocnemius. Červená vlákna jsou silná vlákna. V sarkoplazmě je hodně myoglobinu a v mitochondriích cytochromu, ale méně myofibril. Pomalý začátek, ale vydrží dlouho. Příklad: zádové svaly, bránice.
 Každý sval má síť krevních cév. Svalové kontrakce podporují průtok krve. V uvolněném, nepracujícím svalu je většina krevních kapilár uzavřena pro průtok krve. Při kontrakci svalu se okamžitě otevřou všechny krevní kapiláry.
Každý sval má síť krevních cév. Svalové kontrakce podporují průtok krve. V uvolněném, nepracujícím svalu je většina krevních kapilár uzavřena pro průtok krve. Při kontrakci svalu se okamžitě otevřou všechny krevní kapiláry.

 Struktura svalu Každý sval je na jednom konci připojen k jedné kosti (počátek svalu) a na druhém konci k druhému (úpon svalu). Sval se dělí na: hlavu, břicho a ocas.
Struktura svalu Každý sval je na jednom konci připojen k jedné kosti (počátek svalu) a na druhém konci k druhému (úpon svalu). Sval se dělí na: hlavu, břicho a ocas.
 Vlákna motorických nervů se přibližují ke každému svalovému vláknu a vlákna senzorických nervů odcházejí Počet nervových zakončení ve svalu závisí na stupni funkční aktivity svalů.
Vlákna motorických nervů se přibližují ke každému svalovému vláknu a vlákna senzorických nervů odcházejí Počet nervových zakončení ve svalu závisí na stupni funkční aktivity svalů.

 Každé svalové vlákno je inervováno nezávisle a je obklopeno sítí hemokapilár, tvořících komplex zvaný myon. Skupina svalových vláken inervovaných jedním motorickým neuronem se nazývá motorická jednotka. Je charakteristické, že svalová vlákna patřící do jedné motorické jednotky neleží vedle sebe, ale jsou umístěna mozaikově mezi vlákny patřícími k jiným jednotkám.
Každé svalové vlákno je inervováno nezávisle a je obklopeno sítí hemokapilár, tvořících komplex zvaný myon. Skupina svalových vláken inervovaných jedním motorickým neuronem se nazývá motorická jednotka. Je charakteristické, že svalová vlákna patřící do jedné motorické jednotky neleží vedle sebe, ale jsou umístěna mozaikově mezi vlákny patřícími k jiným jednotkám.


 Šlacha je hustý vláknitý provazec pojivové tkáně, který spojuje sval s kostrou nebo se ke kostře připojuje.
Šlacha je hustý vláknitý provazec pojivové tkáně, který spojuje sval s kostrou nebo se ke kostře připojuje.
 peritenonium kolagenová vlákna typu IV endotenonium Kolagenová vlákna šlachy, propletená s kolagenovými vlákny periostu, jsou vetkána do základní hmoty kostní tkáně a vytvářejí na kostech vyvýšeniny, hrbolky, hrbolky, prohlubně a prohlubně.
peritenonium kolagenová vlákna typu IV endotenonium Kolagenová vlákna šlachy, propletená s kolagenovými vlákny periostu, jsou vetkána do základní hmoty kostní tkáně a vytvářejí na kostech vyvýšeniny, hrbolky, hrbolky, prohlubně a prohlubně.
 Fascie jsou kolagenová vlákna pojivové tkáně s malou příměsí elastických vláken Povrchová temporální fascie Hluboká fascie stehna
Fascie jsou kolagenová vlákna pojivové tkáně s malou příměsí elastických vláken Povrchová temporální fascie Hluboká fascie stehna
 1. 2. 3. 4. 5. Fascie odděluje svaly od kůže a eliminuje posuny kůže při pohybech kontrahujících svalů. Fascie zachovává sílu svalové kontrakce tím, že eliminuje tření mezi svaly během kontrakce. Fascie protahují velké žíly pod napětím, v důsledku čehož je do těchto žil „nasávána“ krev z periferie. Fascie jsou důležité jako bariéry, které brání šíření infekce a nádorů. Během operací pomáhá fascie určit umístění svalů, krevních cév a vnitřností.
1. 2. 3. 4. 5. Fascie odděluje svaly od kůže a eliminuje posuny kůže při pohybech kontrahujících svalů. Fascie zachovává sílu svalové kontrakce tím, že eliminuje tření mezi svaly během kontrakce. Fascie protahují velké žíly pod napětím, v důsledku čehož je do těchto žil „nasávána“ krev z periferie. Fascie jsou důležité jako bariéry, které brání šíření infekce a nádorů. Během operací pomáhá fascie určit umístění svalů, krevních cév a vnitřností.
 Klasifikace svalů Kosterní svaly se liší tvarem, strukturou, polohou vzhledem k osám kloubů atd., a proto jsou klasifikovány různě.
Klasifikace svalů Kosterní svaly se liší tvarem, strukturou, polohou vzhledem k osám kloubů atd., a proto jsou klasifikovány různě.

 III. Podle funkčních vlastností Statické (silné) - krátké břicho a dlouhá šlacha. Svaly pracují s větší silou, ale s menším rozsahem pohybu. Dynamické (obratné) – dlouhé svalové snopce, krátké šlachy. Svaly pracují s menší silou, ale produkují větší pohyby
III. Podle funkčních vlastností Statické (silné) - krátké břicho a dlouhá šlacha. Svaly pracují s větší silou, ale s menším rozsahem pohybu. Dynamické (obratné) – dlouhé svalové snopce, krátké šlachy. Svaly pracují s menší silou, ale produkují větší pohyby
 Pomocný aparát svalů Kosterní svaly mají pomocný aparát, který usnadňuje jejich činnost. n n n Fascia; Osteofasciální pouzdra; synoviální burzy; Synoviální šlachové pochvy; Svalové bloky; Sezamské kosti.
Pomocný aparát svalů Kosterní svaly mají pomocný aparát, který usnadňuje jejich činnost. n n n Fascia; Osteofasciální pouzdra; synoviální burzy; Synoviální šlachové pochvy; Svalové bloky; Sezamské kosti.
 Anomálie svalového vývoje jsou velmi časté a dělí se do tří skupin: 1. Absence jakéhokoli svalu; 2. Přítomnost dalšího svalu, který v přírodě neexistuje. 3. Další snopce stávajícího svalu.
Anomálie svalového vývoje jsou velmi časté a dělí se do tří skupin: 1. Absence jakéhokoli svalu; 2. Přítomnost dalšího svalu, který v přírodě neexistuje. 3. Další snopce stávajícího svalu.
 Vývojové vady: Nevyvinutí m. sternocleidomastoideus - Torticollis, Nevyvinutí bránice. Příčina diafragmatické kýly. Nevyvinutí deltového a trapézového svalu – Deformace pletence a ramene
Vývojové vady: Nevyvinutí m. sternocleidomastoideus - Torticollis, Nevyvinutí bránice. Příčina diafragmatické kýly. Nevyvinutí deltového a trapézového svalu – Deformace pletence a ramene


 I. Tvar: Vřetenovitý; Ve tvaru stuhy; Plochá široká; Zoubkovaný; Dlouho; n n n Čtverec; Trojúhelníkový; Kolo; deltový sval; Soleus atd.
I. Tvar: Vřetenovitý; Ve tvaru stuhy; Plochá široká; Zoubkovaný; Dlouho; n n n Čtverec; Trojúhelníkový; Kolo; deltový sval; Soleus atd.



 II. Ve směru svalových vláken S rovnými paralelními vlákny; S příčnými; S kruhovým; Zpeřené: A. Jednozpeřené; bipinnate; C. Vícezpeřené. B.
II. Ve směru svalových vláken S rovnými paralelními vlákny; S příčnými; S kruhovým; Zpeřené: A. Jednozpeřené; bipinnate; C. Vícezpeřené. B.

 IV. Podle funkce: Adductors; Převaděče; Ohýbání; Extensor; Pronátory; n n Podpěry oblouku; Namáhání; Svaly jsou synergisty; Svaly jsou antagonisté.
IV. Podle funkce: Adductors; Převaděče; Ohýbání; Extensor; Pronátory; n n Podpěry oblouku; Namáhání; Svaly jsou synergisty; Svaly jsou antagonisté.
 V. Ve vztahu ke spoji: Jednokloubový; Dvoukloubový; Vícekloubový.
V. Ve vztahu ke spoji: Jednokloubový; Dvoukloubový; Vícekloubový.
Svalová soustava se vyvíjí souběžně s vývojem nervové soustavy jako jejího výkonného orgánu. Ve fylogenezi se objevuje na základě vlastností kontraktility buněčných elementů nejjednodušších mnohobuněčných organismů, které reagují na podráždění. Zpočátku se v těle zvířete objevuje hladká svalová tkáň, která si stále dobře zachovává svou buněčnou strukturu a je spojena s nervovým systémem prostřednictvím dvou neuronů. (Eferentní komunikace s centry nervového systému probíhá přes periferní neuron.) Se zdokonalováním forem translačního pohybu a s tím spojených podpůrných prvků těla - kostry se objevuje kosterní (somatická) svalová tkáň, která již má přímé eferentní spojení s centrálním nervovým systémem.
Pokud u koelenterátních zvířat nejsou svalové elementy ještě izolované, ale jsou reprezentovány speciálními svalovými výběžky epiteliálních buněk, které se diferencují na podélně umístěné (v ektodermu) a prstencové (v endodermu), pak jsou u červů izolovány, již nejsou spojeny s epitelem a tvoří muskulokutánní vak.
U strunatců, pohyblivějších zvířat, se všechny svaly dělí na somatické a viscerální. U dospělých jsou somatické svaly příčně pruhované. Jsou představovány podélnou vrstvou párových laterálních svalů, rozdělených membránami na samostatné metamery (segmenty). Lancelet má stále hladkou svalovou tkáň ve svých cévách a střevech. U obratlovců, stejně jako u lanceletu, jsou svaly rozděleny na somatické a hladké, liší se nejen strukturou, ale také povahou nervových spojení.
Čím složitější je organismus obratlovců ve fylogenezi, tím více kosterní svaly ztrácejí svou metamerii, tím jsou diferencovanější, dělí se nejprve na dorzální a ventrální vrstvy a poté na samostatné vrstvy, úseky, svalové skupiny a svaly.
U suchozemských obratlovců jsou již všechny kosterní svaly rozděleny do skupin: svaly trupu, hlavy a končetin. Hladká svalová tkáň stále zůstává především ve stěnách cév a vnitřních orgánů. Bez ohledu na to, jak složitý je kosterní sval jako orgán, je spojen s určitými segmenty mozku.
Toto spojení vzniká od prvních okamžiků objevení se svalových elementů a není přerušeno během fylogenetických přeměn. V tomto ohledu bylo zjištěno, že skupina svalových vláken přijímajících nervová zakončení (nervové plaky) z jedné nervové buňky je „sval jednotka“. Z kterého segmentu se sval v zárodku vyvíjí, ze stejného segmentu je v budoucnu inervován. Pokud se sval během vývoje pohne, následuje nerv (ukázkovým příkladem toho je bránice umístěná za hrudním provazcem). Pokud se sval diferencuje na samostatné části, nerv podstoupí stejnou diferenciaci; splyne-li sval, splynou i nervy. Redukce svalu vede k redukci nervového spojení. Takové neměnné spojení umožňuje identifikovat cesty vývoje svalů v embryogenezi a fylogenezi a umožňuje stanovit, pomocí kterých segmentů embrya bylo vytvořeno.
Obrázek 91. Schéma zevního svalstva obratlovců
Během procesu individuálního vývoje (v ontogenezi) se svaly vyvíjejí z několika zdrojů mezodermu; 1) kvůli materiálu mezodermových myotomů; 2) z nesegmentovaného mezodermu v oblasti žáber. Proto se svaly, které se vyvíjejí díky mezenchymu žaberního aparátu, nazývají viscerální. Zpočátku je v embryu analáž svalových prvků, stejně jako ve fylogenezi, segmentována a diferencována na samostatné metamery, oddělené přepážkami pojivové tkáně.
V ontogenezi se hlavní etapy opakují, zejména v raném období fylogeneze (obr. 91). Zpočátku je laterální sval položen podél těla jako podélná párová šňůra, segmentovaná přepážkami pojivové tkáně na metamerně umístěné myotomy a každý myotom přijímá nerv určitého nervového segmentu. Poté se objeví podélná přepážka pojivové tkáně, která rozděluje myotom na dorzální a ventrální část. Následně se na místě této podélné přepážky vyvinou příčné výběžky obratlů a žeber. Ve stejné sekvenci dochází k rozdělení nervových větví, přičemž se zachovají spojení se stejnými nervovými segmenty. Díky materiálu dorzálních částí myotomů se následně vyvíjejí svaly páteře a díky ventrálním partiím se vyvíjejí svaly laterální hrudní a břišní stěny.
O něco později svalové snopce vrůstají do končetinových pupenů z dorzálních a ventrálních úseků myotomů (z pěti myotomů na prsních končetinách a z osmi na pánevních končetinách). S diferenciací svalových vrstev na jednotlivé svaly se kosterní svaly stávají ještě komplexnějšími, zejména s rozestupem svalů končetin, které jsou také inervovány převážně ventrálními větvemi míšních nervů.
Svaly hlavy a některé svaly krku se vyvíjejí z nesegmentovaného mezodermu v oblasti prvních pěti větevních oblouků. Z rudimentů prvního branchiálního oblouku se vyvíjejí žvýkací svaly spojené s V párem hlavových nervů, z druhého - obličejové svaly spojené s VII párem hlavových nervů.
Svalová tkáň se díky své schopnosti stahovat se během procesu fylogeneze zformovala do samostatných orgánů - svalů - pásových motorů se silou schopnou zvedat těžké váhy, podpírat tělesnou hmotnost na nohou, dodávat údery, vyvíjet vysokou rychlost při pohybu , atd.
Svaly při kontrakci nejen způsobují pohyb (provádějí dynamickou práci), ale také zajišťují svalový tonus, posilují klouby v určitém úhlu kombinace se stacionárním tělem, udržují určité držení těla (provádějí statickou práci). Statická práce, zejména u velkých (těžkých) kopytníků, je extrémně náročná.
Určitý svalový tonus plní i antigravitační funkci. Spolu se šlachami, fasciemi a vazy kostry mají svaly elastické vlastnosti, díky nimž jsou spolehlivými tlumiči nárazů, dodávají končetinám větší pružnost a fungují také jako mikropumpy, které podporují odvod krve a lymfy z orgánů (N. I. Ariinchin, 1987).
U každého druhu zvířete v procesu evoluce svaly, vyvíjející se, určují své limity síly, rychlosti pohybu, vytrvalosti a také hmotnosti, která je přímo závislá na druhu, věku, pohlaví, plemeni zvířete. a jejich výcvik. Svalová hmota tvoří přibližně 42-47 % celkové tělesné hmotnosti u skotu a koní, 34 % u ovcí a 31 % u prasat. Pouze práce (trénink) svalů pomáhá zvyšovat jejich hmotu jak zvětšením průměru svalových vláken (hypertrofie), tak zvýšením jejich počtu (hyperplazie).
Práce svalů je úzce spjata s orgánem rovnováhy a do značné míry i s ostatními smyslovými orgány. Díky tomuto spojení svaly poskytují tělu rovnováhu, přesnost pohybů a sílu. Svaly jsou výkonným generátorem přeměny chemické energie potravy na tepelnou energii, energie pružných deformací na mechanickou energii, která se využívá k pohybu krve cévami a vybuzení aferentních impulsů vysílaných do mozku, jakož i k přechodu do tepelné energie.
Přednáška z biologie na téma "Fylogeneze pohybového a nervového systému" Fylogeneze svalového systému: zákonitosti vývoje
Domácí sušenky se smetanou
Thajský král Bhumibol Adulyadej: božský král s kamerou
Prasátko - biblické otázky
Recept: Cuketové palačinky - Se sýrem - velmi jemné a chutné