– ряд ядерных структур, занимающих центральное место в стволовой части мозга. Морфологическое строение ретикулярной формации очень напоминает сеточку, и именно на этот признак в первую очередь обратил внимание немецкий анатом Отто Дейтерс. Именно по этому признаку он и назвал эту структуру (лат. reticulum - сеточка, formatio - образование). Согласен! «Сетчатое образование» — звучит не так круто 🙂
Так как ретикулярная формации проходит через весь ствол головного мозга, то анатомически её можно было бы разделить на отделы продолговатого мозга, варолиева моста и среднего мозга, но т. к. отдельные части этой структуры заняты общим для них делом, то ее рассматривают, как единую структуру.
Для понимания приведу сравнение… Если вы когда-нибудь бывали на концертах или хотя бы смотрели их по телевизору, то наверняка замечали огромные панели с кучей кнопочек, тумблеров, переключателей и т. д. С помощью этих панелей оператор настраивает качество звука, приглушая одни частоты и повышая другие, а так же цветность, яркость, контраст и т. д. Так вот ретикулярная формация занимается именно этим. Т. е. она получает сигналы абсолютно от всех нисходящих и восходящих путей, перерабатывает их, вырабатывает новые координирующие сигналы и выдает их по назначению, обеспечивая нам нормальное восприятие.
Некоторые нейроны Р. ф. проявляют фоновую активность, разряжаясь 5-10 раз в секунду. Эти нервные центры оказывают влияния на кору головного мозга, постоянно поддерживая в нас сознание. При разрушении этих центров у животных наступает .
Для сравнения поясню. Ретикулярная формация поддерживает в нас сознание так же, как огонь в костре поддерживает кипение воды в котле. Стоит потушить огонь — и кипящая вода, так же как и сознание, перестает проявлять активность. Следовательно, в Р. ф. находятся одни из центров сна и бодрствования.
С корой головного мозга вообще связи особенные. Понятно, что Р.Ф. отвечает за и возбуждения в коре мозга. Кора мозга, в свою очередь, также оказывает тормозные и возбуждающие влияния на ретикулярную формацию. Образуя замкнутые нейронные связи, эти две системы взаимно регулируют друг друга и уравновешивают свои влияния.
Выводы
Функции ретикулярной формации изучены не достаточно ввиду высокой степени сложности организации данной структуры, но имеющихся данных достаточно для следующих выводов:
Оказывает влияние на уровень сознания путем взаимодействия в кору мозга. Участвует в цикле в придание эмоциональной окраски сенсорным сигналам в т. ч. болевым, путем проведения афферентной информации к лимбической системе. Путем взаимного координирования афферентных и эфферентных систем участвует в формировании жизненно важных рефлексов. Также принимает участие в вегетативных функциях организма и как важная составляющая двигательных центров ствола мозга.
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
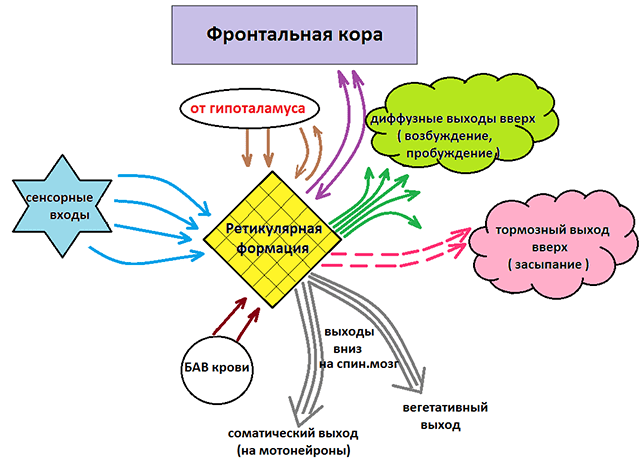
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Лекция 6.
Ретикулярная формация — это комплекс нейронов ствола головного мозга и частично спинного мозга, который имеет обширные связи с различными нервными центрами, корой головного мозга и между собой. Ретикулярная формация представлена рассеянными клетками в покрышке ствола мозга и в спинном мозге.
Ряд клеток ретикулярной формации в стволе мозга являются жизненно важными центрами:
1. дыхательным (центр вдоха и выдоха) — в продолговатом мозге;
2. сосудодвигательным - в продолговатом мозге;
3. центром координации взора (ядра Кахаля и Даркшевича) — в среднем мозге;
4. центром терморегуляций — в промежуточном мозге;
5. центром голода и насыщения — в промежуточном мозге.
Ретикулярная формация выполняет следующие функции:
Обеспечение сегментарных рефлексов: рассеянные клетки являются
вставочными нейронами спинного мозга и ствола головного мозга
(рефлекс глотания);
Поддержание тонуса скелетной мускулатуры: клетки ядер ретикулярной формации посылают тонические импульсы на двигательные ядра черепных нервов и двигательные ядра передних рогов спинного мозга;
Обеспечение тонической активности ядер ствола головного мозга и
коры полушарий, что необходимо для дальнейшего проведения и
анализа нервных импульсов;
Коррекция при проведении нервных импульсов: благодаря ретикулярной формации импульсы могут либо существенно усиливаться, либо существенно ослабляться в зависимости от состояния нервной системы;
Активное влияние на высшие центры коры больших полушарий, что
приводит к либо снижению тонуса коры, апатии и наступлению сна,
либо к повышению работоспособности, эйфории;
Участие в регуляции сердечной деятельности, дыхания, тонуса сосудов,
секреции желез и других вегетативных функций (центры ствола мозга);
Участие в регуляции сна и бодрствования: голубое пятно, ядра шва —
проецируются на ромбовидную ямку;
Обеспечение сочетанного поворота головы и глаз: ядра Кахаля и
Даркшевича.
Основным нисходящим трактом ретикулярной формации является ретикулоспинальный, который проходит по стволу к нейронам двигательных ядер передних рогов спинного мозга и двигательных ядер черепных нервов, а также к вставочным нейронам вегетативной нервной системы.
От ретикулярных ядер зрительного бугра к различным областям коры больших полушарий идут таламо-корковые волокна: они заканчиваются во всех слоях коры большого мозга, осуществляя активацию коры, необходимую для восприятия специфических раздражений.
⇐ Предыдущая22232425262728293031
| Защита персональных данных |
Не нашли то, что искали? Воспользуйтесь поиском:
Читайте также:
Главная → Физиология → Механизм регуряции физиологических процессов ->
Функции ретикулярной формации
Ретикулярная формация
ствола мозга рассматривается как один из важных интегративных аппаратов мозга.
К собственно интегративных функций ретикулярной формации относятся:
- контроль над состояниями сна и бодрствования
- мышечный (фазный и тонический) контроль
- обработка информационных сигналов окружающей и внутренней среды организма, которые поступают по разным каналам
Ретикулярная формация объединяет различные участки ствола мозга (ретикулярную формацию продолговатого мозга, варолиева моста и среднего мозга). В функциональном отношении в ретикулярной формации разных отделов мозга есть много общего, поэтому целесообразно рассматривать ее как единую структуру. Ретикулярная формация представляет собой диффузное накопление клеток разного вида и величины, которые разделены многими волокнами. Кроме этого, в середине ретикулярной формации выделяют около 40 ядер и пидьядер.
Ретикулярная формация мозга: строение и функции
Нейроны ретикулярной формации имеют широко разветвленные дендриты и продолговатые аксоны, часть которых делится Т-образно (один отросток направлен вниз, образуя ретикулярный-спинальный путь, а второй — в верхние отделы головного мозга).
В ретикулярной формации сходится большое количество афферентных путей из других мозговых структур: из коры большого мозга — коллатерали кортико-спинальных (пирамидных) путей, из мозжечка и других структур, а также коллатеральные волокна, которые подходят через ствол мозга, волокна сенсорных систем (зрительные, слуховые и т.д.). Все они заканчиваются синапсами на нейронах ретикулярной формации. Так, благодаря такой организации ретикулярная формация приспособлена к объединению влияний из различных структур мозга и способна влиять на них, то есть выполнять интегративные функции в деятельности ЦНС, определяя в значительной мере общий уровень ее активности.
Свойства ретикулярных нейронов. Нейроны ретикулярной формации способны к устойчивой фоновой импульсной активности. Большинство из них постоянно генерирует разряды частотой 5-10 Гц. Причиной такой постоянной фоновой активности ретикулярных нейронов являются: во-первых, массивная конвергенция различных афферентных влияний (от рецепторов кожных, мышечных, висцеральных, глаза, уши и др.)., А также воздействий из мозжечка, коры большого мозга, вестибулярных ядер и других мозговых структур на один и тот же ретикулярный нейрон. При этом зачастую в ответ на это возникает возбуждение. Во-вторых, активность ретикулярного нейрона может быть изменена гуморальными факторами (адреналин, ацетилхолин, напряжение С02 в крови, гипоксия и др.).. Эти непрерывные импульсы и химические вещества, содержащиеся в крови, поддерживают деполяризацию мембран ретикулярных нейронов, их способность к устойчивой импульсной активности. В связи с этим ретикулярная формация тоже оказывает на другие мозговые структуры постоянный тонический влияние.
Характерной особенностью ретикулярной формации также высокая чувствительность ее нейронов в различных физиологически активных веществ. Благодаря этому деятельность ретикулярных нейронов может быть сравнительно легко блокирована фармакологическими препаратами, которые связываются с циторецепторамы мембран этих нейронов. Особенно активными в этом отношении соединения барбитуровой кислоты (барбитураты), аминазин и другие лекарственные препараты, которые широко применяются в медицинской практике.
Характер неспецифических влияний ретикулярной формации. Ретикулярная формация ствола мозга участвует в регуляции вегетативных функций организма. Однако еще в 1946 г. американский нейрофизиолог Н. W. Megoun и его сотрудники обнаружили, что ретикулярная формация имеет непосредственное отношение к регуляции соматической рефлекторной деятельности. Было доказано, что ретикулярная формация оказывает диффузное неспецифическое, нисходящее и восходящее влияние на другие мозговые структуры.
Нисходящее влияние.
При раздражении ретикулярной формации заднего мозга (особенно гигантоклеточной ядра продолговатого мозга и ретикулярного ядра моста, где принимают начало ретикулоспинальному пути), возникает торможение всех спинальных двигательных центров (сгибательных и разгибательных). Это торможение очень глубокое и продолжительное. Такое положение в естественных условиях может наблюдаться при глубоком сне.
Наряду с диффузными тормозящими влияниями, при раздражении определенных участков ретикулярной формации выявляется диффузное влияние, которое облегчает деятельность спинальной двигательной системы.
Ретикулярная формация играет важную роль в регуляции деятельности мышечных веретен, изменяя частоту разрядов, поступающие гамма-эфферентными волокнами к мышцам. Таким образом модулируется обратная импульсация в них.
Восходящий влияние. Исследования Н. W. Megoun, G. Moruzzi (1949) показали, что раздражение ретикулярной формации (заднего, среднего и промежуточного мозга) сказывается на деятельности высших отделов головного мозга, в частности коры большого мозга, обеспечивая переход ее в активное состояние. Это положение подтверждается данными многочисленными экспериментальными исследованиями и клиническими наблюдениями. Так, если животное находится в состоянии сна, то прямое раздражение ретикулярной формации (особенно варолиева моста) через введенные в эти структуры электроды вызывает поведенческую реакцию пробуждения животного. При этом на ЭЭГ возникает характерное изображение — изменение альфа-ритма бета-ритмом, т.е. фиксируется реакция десинхронизации или активизации. Указанная реакция не ограничивается определенным участком коры большого мозга, а охватывает большие ее массивы, т.е. носит генерализованный характер. При разрушении ретикулярной формации или выключении ее восходящих связей с корой большого мозга животное впадает в сноподобное состояние, не реагирует на световые и обонятельные раздражители, фактически не вступает в контакт с внешним миром. То есть конечный мозг прекращает активно функционировать.
Таким образом, ретикулярная формация ствола головного мозга выполняет функции восходящей активирующей системы мозга, которая поддерживает на высоком уровне возбудимость нейронов коры большого мозга.
Кроме ретикулярной формации ствола мозга, в восходящую активирующую систему головного мозга входят также неспецифические ядра таламуса, задний гипоталамус, лимбических структуры. Являясь важным интегративным центром, ретикулярная формация, в свою очередь, является частью более глобальных интеграционных систем мозга, которые включают гипоталамо-лимбические и неокортикальные структуры. Именно во взаимодействии с ними и формируется целесообразное поведение, направленное на приспособление организма к меняющимся условиям внешней и внутренней среды.
Одним из основных проявлений повреждения ретикулярных структур у человека является потеря сознания. Она бывает при черепно-мозговых травмах, нарушении мозгового кровообращения, опухолях и инфекционных процессах в стволе мозга. Длительность состояния обморока зависит от характера и выраженности нарушений функции ретикулярной активизирующей системы и колеблется от нескольких секунд до многих месяцев. Дисфункция восходящих ретикулярных влияний проявляется тоже потерей бодрости, постоянной патологической сонливостью или частыми приступами засыпания (пароксизмальная гиперсомия), беспокойным ночным сном. Наблюдаются также нарушения (чаще повышении) мышечного тонуса, различные вегетативные изменения, эмоционально-психические расстройства и др.
категории раздела
Тема 13. Ретикулярная формация.
Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур — наличие особых ретикулярных нейронов и отличительный характер связей.
Рис. 1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно , находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество — норадреналин , который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин , который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича , относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся.
Что такое ретикулярная формация
В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
 |
1-6 - медианная зона РФ: 1-4- ядра шва (1 - бледное, 2 - темное, 3 - большое, 4- мостовое), 5 - верхнее центральное, 6 - дорсальное ядро шва, 7-13 - медиальная зона РФ: 7 - ретикулярное парамедианное, 8 - гигантоклеточное, 9 - ретикулярное ядро покрышки моста, 10, 11 - каудальное (10) и оральное (11) ядра моста, 12 - дорсальное покрышечное ядро (Гуддена), 13 - клиновидное ядро, 14- I5-латеральная зона РФ: 14 - центральное ретикулярное ядро продолговатого мозга, 15 - латеральное ретикулярное ядро, 16, 17 - медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 - компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Поиск Лекций
IV. Ретикулярная формация
Ретикулярная формация – протяженная структура в стволе мозга – важная интегративная область неспецифической системы. Первые описания ретикулярной формации (РФ) ствола мозга были сделаны немецкими морфологами: в 1861 г. К. Рейхертом (Reichert K., 1811-1883) и в 1863 г. О. Дейтерсом (Deiters O., 1834-1863); из отечественных исследователей большой вклад в ее изучение внес В.М. Бехтерев. РФ — это совокупность нервных клеток и их отростков, расположенных в покрышке всех уровней ствола между ядрами черепных нервов, оливами, проходящими здесь афферентными и эфферентными проводящими путями (рисунок 17). К ретикулярной формации иногда от носят и некоторые медиальные структуры промежуточного мозга, в том числе медиальные ядра таламуса.
Клетки РФ различны по форме и величине, длине аксонов, расположены преимущественно диффузно, местами образуют скопления — ядра, которые обеспечивают интеграцию импульсов, поступающих от расположенных поблизости черепных ядер или проникающих сюда по коллатералям от проходящих через ствол афферентных и эфферентных проводящих путей. Среди связей ретикулярной формации ствола мозга важнейшими можно считать корково- ретикулярные, спинно-ретикулярные пути, связи между РФ ствола с образованиями промежуточного мозга и стриопаллидарной системой, мозжечково-ретикулярные пути. Отростки клеток РФ формируют афферентные и эфферентные связи между содержащимися в покрышке ствола ядрами черепных нервов и проекционными проводящими путями, входящими в состав покрышки ствола. По коллатералям от проходящих через ствол мозга афферентным путям РФ получает «подзаряжающие» ее импульсы и выполняет при этом функции аккумулятора и генератора энергии. Следует отметить и высокую чувствительность РФ к гуморальным факторам, в том числе к гормонам, лекарственным средствам, молекулы которых достигают ее гематогенным путем.

Рис.17. Ретикулярная формация.
Нейроны ретикулярной формации собраны в ядра, выполняющие специфические функции, и посылают отростки в большинство областей мозговой коры. Различают восходящую ретикулярную систему (слева), вызывающую активацию коры, и нисходящую ретикулярную систему (справа), главным образом регулирующую постуральный тонус (поддержание позы) благодаря тормозному и облегчающему влиянию на двигательные пути, спускающиеся из моторной коры в спинной мозг
К восходящей активирующей системе относятся ядра ретикулярной формации, расположенные, главным образом, на уровне среднего мозга, к которым подходят коллатерали от восходящих чувствительных систем. Возникающие в этих ядрах нервные импульсы по полисинаптическим проводящим путям, проходя через интраламинарные ядра таламуса, субталамические ядра к коре больших полушарий, оказывают на нее активирующее влияние. Восходящие влияния неспецифической активирующей ретикулярной системы имеют большое значение в регуляции тонуса коры больших полушарий, а также в регуляции процессов сна и бодрствования.
В случаях поражения активирующих структур ретикулярной формации, а также при нарушении ее связей с корой больших полушарий возникает снижение уровня сознания, активности психической деятельности, в частности когнитивных функций, двигательной активности. Возможны проявления оглушенности, общей и речевой гипокинезии, акинетического мутизма, сопора, комы, вегетативного состояния.
В составе РФ имеются отдельные территории, получившие в процессе эволюции элементы специализации — вазомоторный центр (депрессорные и прессорные его зоны), дыхательный центр (экспираторный и инспираторный), рвотный центр. РФ содержит структуры, влияющие на соматопсиховегетативную интеграцию . РФ обеспечивает поддержание витальных рефлекторных функций — дыхания и сердечно-сосудистой деятельности, принимает участие в формировании таких сложных двигательных актов, как кашель, чиханье, жевание, рвота, сочетанная работа речедвигательного аппарата, общей двигательной активности.
Нисходящие влияния РФ на спинной мозг сказываются прежде всего на состоянии мышечного тонуса и могут быть активирующими или понижающими мышечный тонус, что важно для формирования двигательных актов. Обычно активация или торможение восходящих и нисходящих влияний РФ осуществляется параллельно. Так, во время сна, для которого характерно торможение восходящих активирующих влияний, происходит торможение и нисходящих неспецифических проекций, что проявляется, в частности, снижением тонуса мышц.
Функции РФ еще не вполне изучены. Считается, что она участвует в ряде процессов:
– регуляция возбудимости коры: уровня осознания стимулов и реакций, ритма сон–бодрствование (восходящая активирующая ретикулярная система);
– придание аффективно–эмоциональной окраски сенсорным стимулам, особенно болевым, за счет передачи афферентной информации в лимбическую систему;
– двигательная регуляция функций, в том числе жизненно важных рефлексов (кровообращения, дыхания, глотания, кашля и чихания), при которых должны взаимно координироваться разные афферентные и эфферентные системы;
– участие в регуляции позных и целенаправленных движений в качестве важного компонента двигательных центров ствола мозга.
V. Мозжечок
Мозжечок располагается под дубликатурой твердой мозговой оболочки, известной как намет мозжечка , который разделяет полость черепа на два неравных пространства — супратенториальное и субтенториальное. В субтенториальном пространстве, дном которого является задняя черепная ямка, помимо мозжечка, находится ствол мозга. Объем мозжечка составляет в среднем 162 см3. Масса его варьирует в пределах 136-169 г.
Мозжечок находится над мостом и продолговатым мозгом. Вместе с верхним и нижним мозговыми парусами он составляет крышу IV желудочка мозга, дном которого является так называемая ромбовидная ямка. Над мозжечком находятся затылочные доли большого мозга, отделенные от него наметом мозжечка.
В мозжечке различают два полушария . Между ними в сагиттальной плоскости над IV желудочком мозга располагается филогенетически наиболее древняя часть мозжечка — его червь . Червь и полушария мозжечка фрагментируются на дольки глубокими поперечными бороздами.
Мозжечок состоит из серого и белого веществ. Серое вещество формирует кору мозжечка и находящиеся в его глубине парные ядра (рисунок 18). Самые крупные из них — зубчатые ядра — расположены в полушариях. В центральной части червя имеются ядра шатра , между ними и зубчатыми ядрами находятся шаровидные и пробковидные ядра .

Рис. 18.Ядра мозжечка.
1 — зубчатое ядро; 2 — пробковидное ядро; 3 — ядро шатра; 4 — шаровидное ядро.

Рис. 19. Сагиттальный срез мозжечка и ствола мозга.
1 — мозжечок; 2 — «древо жизни»; 3 — передний мозговой парус; 4 — пластинка четверохолмия; 5 — водопровод мозга; 6 — ножка мозга; 7 — мост; 8 — IV желудочек, его сосудистое сплетение и шатер; 9 — продолговатый мозг.
Ввиду того, что кора покрывает всю поверхность мозжечка и проникает в глубину его борозд, на сагиттальном разрезе мозжечка ткань его имеет рисунок листа, прожилки которого образованы белым веществом (рисунок 19), составляющим так называемое древо жизни мозжечка . В основании древа жизни находится клиновидная выемка, являющаяся верхней частью полости IV желудочка; края этой выемки образуют его шатер. Крышей шатра служит червь мозжечка, а переднюю и заднюю его стенки составляют тонкие мозговые пластинки, известные под названием переднего и заднего мозговых парусов.
В кору мозжечка импульсы поступают по проникающим в нее из белого вещества мшистым и ползучим волокнам, составляющим афферентные пути мозжечка.
Функции ретикулярной формацииПо мшистым волокнам импульсы, поступающие из спинного мозга, вестибулярных ядер и ядер моста, передаются на клетки зернистого слоя коры. Аксоны этих клеток вместе с ползучими волокнами, проходящими через зернистый слой транзитом и несущими в мозжечок импульсы от нижних олив, доходят до поверхностного, молекулярного слоя мозжечка. Здесь аксоны клеток зернистого слоя и ползучие волокна Т-образно делятся, причем в молекулярном слое их разветвления принимают направление, продольное поверхности мозжечка. Импульсы, достигшие молекулярного слоя коры, пройдя через синаптические контакты, попадают на располагающиеся здесь же разветвления дендритов клеток Пуркинье. Далее они следуют по дендритам клеток Пуркинье к их телам, расположенным на границе молекулярного и зернистого слоев. Затем по аксонам тех же клеток, пересекающих зернистый слой, проникают в глубину белого вещества. Заканчиваются аксоны клеток Пуркинье в ядрах мозжечка. Главным образом в зубчатом ядре. Эфферентные импульсы, идущие от мозжечка по аксонам клеток, составляющих его ядра и принимающих участие в формировании мозжечковых ножек, покидают мозжечок. |
Мозжечок имеет три пары ножек: нижнюю, среднюю и верхнюю. Нижняя ножка связывает его с продолговатым мозгом, средняя — с мостом, верхняя — со средним мозгом. Ножки мозга составляют проводящие пути, несущие импульсы к мозжечку и от него.
Червь мозжечка обеспечивает стабилизацию центра тяжести тела, его равновесие, устойчивость, регуляцию тонуса реципрокных мышечных групп, главным образом шеи и туловища, и возникновение при этом физиологических мозжечковых синергий, стабилизирующих равновесие тела.
Для успешного поддержания равновесия тела мозжечок постоянно получает информацию, проходящую по спиноцеребеллярным путям от проприоцепторов различных частей тела, а также от вестибулярных ядер, нижних олив, ретикулярной формации и других образований, участвующих в контроле за положением частей тела в пространстве. Большинство афферентных путей, идущих к мозжечку, проходит через нижнюю мозжечковую ножку, часть их расположена в верхней мозжечковой ножке.
Через свои средние ножки мозжечок получает импульсы из коры больших полушарий мозга. Эти импульсы проходят по корково-мостомозжечковым путям.
Часть импульсов, возникших в коре больших полушарий мозга, достигает противоположного полушария мозжечка, принося информацию не о произведенном, а лишь о намечаемом к выполнению активном движении. Получив такую информацию, мозжечок моментально высылает импульсы, корригирующие произвольные движения, главным образом, путем погашения инерции и наиболее рациональной регуляции тонуса реципрокных мышц— мышц-агонистов и антагонистов. В результате создается своеобразная эйметрия,делающая произвольные движения четкими, отточенными, лишенными нецелесообразных компонентов.
Пути, выходящие из мозжечка, состоят из аксонов клеток, тела которых формируют его ядра. Большинство эфферентных путей, в том числе пути, идущие от зубчатых ядер, покидают мозжечок через его верхнюю ножку. На уровне нижних бугров четверохолмия совершается перекрест эфферентных мозжечковых путей (перекрест верхних мозжечковых ножек Вернекинга). После перекреста каждый из них достигает красных ядер противоположной стороны среднего мозга. В красных ядрах мозжечковые импульсы переключаются на следующий нейрон и дальше движутся по аксонам клеток, тела которых заложены в красных ядрах. Эти аксоны формируются в красноядерно-спинномозговые проводящие пути , которые вскоре после выхода из красных ядер подвергаются перекресту (перекрест покрышки или перекрест Фореля), после чего спускаются в спинной мозг. В спинном мозге красноядерноспинномозговые пути располагаются в боковых канатиках; составляющие их волокна заканчиваются у клеток передних рогов спинного мозга.
Из ядер червя мозжечка эфферентные пути идут в основном через нижнюю мозжечковую ножку к ретикулярной формации ствола мозга и вестибулярным ядрам. Отсюда по ретикулоспинномозговым и вестибулоспинномозговым путям, проходящим по передним канатикам спинного мозга, они также достигают клеток передних рогов. Часть импульсов, идущих от мозжечка, пройдя через вестибулярные ядра, попадает в медиальный продольный пучок, доходит до ядер III, IV и VI черепных нервов, обеспечивающих движения глазных яблок, и оказывает влияние на их функцию.
Таким образом:
1. Каждая половина мозжечка получает импульсы в основном а) из гомолатеральной половины тела, б) из противоположного полушария мозга, имеющего кортико-спинальные связи с той же половиной тела.
2. От каждой половины мозжечка эфферентные импульсы направляются к клеткам передних рогов гомолатеральной половины спинного мозга и к ядрам черепных нервов, обеспечивающих движения глазных яблок.
Такой характер мозжечковых связей позволяет понять, почему при поражении одной половины мозжечка мозжечковые расстройства возникают преимущественно в той же, т.е. гомолатеральной, половине тела. Это особенно отчетливо проявляется при поражении полушарий мозжечка.
©2015-2018 poisk-ru.ru
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Нарушение авторских прав и Нарушение персональных данных
Ретикулярная формация
Термин «ретикулярная формация» (англ. ret – сеть) был введен впервые Дейтерсом более 100 лет назад. Ретикулярная формация (РФ) располагается в центральной части мозгового ствола, заходя ростральным концом в таламус, а каудальным – в спинной мозг. Благодаря наличию сетевых связей почти со всеми структурами центральной нервной системы она получила название ретикулярной, или сетевой, формации.
Различные по форме и величине нейроны РФ имеют длинные дендриты и короткий аксон, хотя встречаются гигантские нейроны с длинными аксонами, образующими, например, руброспинальный и ретикулоспинальный тракты. На одной нервной клетке может заканчиваться до 40 000 синапсов, что указывает на широкие межнейрональные связи в пределах РФ. В ней был выделен целый ряд ядер и ядерных групп, отличающихся как в структурном отношении, так и выполняемыми ими функциями.
Ретикулярная формация образует многочисленные как афферентные пути: спиноретикулярный, церебеллоретикулярный, корково‑подкорково‑ретикулярный (от коры, базальных ганглиев, гипоталамуса), от структур каждого уровня ствола мозга (от среднего мозга, варолиева моста, продолговатого мозга), так и эфферентные: нисходящие ретикулоспинальные, ретикулокорково‑подкорковые, ретикуломозжечковые, а также пути к другим структурам ствола мозга.
Ретикулярная формация оказывает генерализованное, тонизирующее, активирующее влияние на передние отделы головного мозга и кору больших полушарий (восходящая активирующая система РФ) и нисходящее, контролирующее деятельность спинного.мозга (нисходящая ретикулоспинальная система), которое может быть как облегчающим на многие функции организма, так и тормозным. Одним из видов тормозного влияния РФ на рефлекторную деятельность спинного мозга является сеченовское торможение, заключающееся в угнетении спинальных рефлексов при раздражении таламической ретикулярной формации кристалликом соли.
Г. Мэгун показал, что локальное электрическое раздражение гигантоклеточного ядра РФ продолговатого мозга вызывает торможение сгибательного и разгибательного рефлексов спинного мозга, а на мотонейроне возникает длительный ТПСП и постсинаптическое торможение по типу гиперполяризации.
Тормозные влияния на сгибательные рефлексы оказывает преимущественно медиальная ретикулярная формация продолговатого мозга, а облегчающие – латеральные зоны РФ моста.
Ретикулярная формация принимает участие в реализации многих функций организма. Так, РФ контролирует двигательную активность, постуральный тонус и фазные движения.
В 1944 г. в США во время эпидемии полиомиелита – заболевания, нарушающего двигательную активность, основные структурные изменения были обнаружены в ретикулярной формации. Это навело американского ученого Г. Мэгуна на мысль об участии РФ в моторной активности. Основными ее структурами, отвечающими за этот вид деятельности, являются ядро Дейтерса продолговатого мозга и красное ядро среднего мозга. Ядро Дейтерса поддерживает тонус альфа‑ и гамма‑мотонейронов спинного мозга, иннервирующих мышцы‑разгибатели, и тормозит альфа‑ и гамма‑мотонейроны мышц‑сгибателей. Красное ядро, напротив, тонизирует альфа‑ и гамма‑мотонейроны мышц‑сгибателей и тормозит альфа‑ и гаммамотонейроны мышц‑разгибателей. Красное ядро оказывает тормозное влияние на ядро Дейтерса, поддерживая равномерный тонус мышц‑разгибателей. Повреждения или перерезка мозга между средним и продолговатым приводит к снятию тормозных влияний со стороны красного ядра на ядро Дейтерса, а значит, и на тонус мышц‑разгибателей, который начинает преобладать над тонусом мышц‑сгибателей и возникает децеребрационная ригидность или повышенный тонус мышц, проявляющийся в сильном сопротивлении растяжению. Такое животное имеет характерную позу тела: запрокинута голова, вытянуты передние и задние конечности. Поставленное на ноги, оно при малейшем толчке падает, так как отсутствует тонкая регуляция позы тела.
Раздражение ретикулярной формации вызывает тремор, спастический тонус.
РФ среднего мозга играет роль в координации сокращений глазных мышц. Получив информацию от верхних бугров четверохолмия, мозжечка, вестибулярных ядер, зрительных областей коры головного мозга, РФ ее интегрирует, что приводит к рефлекторным изменениям работы глазодвигательного аппарата, особенно при внезапном появлении движущихся объектов, изменении положения головы и глаз.
Ретикулярная формация регулирует вегетативные функции, в реализации которых принимают участие так называемые стартовые нейроны РФ, запускающие процесс возбуждения внутри определенной группы нейронов, отвечающих за дыхательные и сосудодвигательные функции. В РФ продолговатого мозга расположены два ядра, одно из них отвечает за вдох, другое – за выдох. Их деятельность контролируется пневмотаксическим центром РФ варолиева моста. Раздражением этих участков РФ можно воспроизвести различные дыхательные акты.
Сосудодвигательный центр расположен в ромбовидной ямке дна четвертого желудочка, входящего в состав РФ. При электрораздражении определенных точек варолиева моста и продолговатого мозга возникают сосудодвигательные реакции.
Ретикулярная формация связана со всеми отделами коры мозга с помощью диффузной неспецифической проекционной афферентной системы, которая, в отличие от специфической, проводит возникшее на периферии возбуждение к коре больших полушарий медленно через последовательно связанные многонейронные системы.
Ретикулярная формация
РФ оказывает активирующее восходящее влияние на кору больших полушарий. Раздражение РФ вызывает «реакцию пробуждения», а на электроэнцефалограмме – десинхронизацию альфа‑ритма и ориентировочный рефлекс.
Перерезка головного мозга ниже РФ вызывает картину бодрствования, выше – сна. РФ регулирует цикл «сон‑бодрствование».
Ретикулярная формация оказывает влияние на сенсорные системы мозга: на остроту слуха, зрения, обонятельные ощущения. Так, повреждение РФ и барбитуровый наркоз приводят к усилению сенсорных импульсов, которые в норме находятся под тормозным, регулирующим влиянием РФ. Восприятие различных ощущений при сосредоточении внимания на каком‑либо другом ощущении, привыкание к повторяющимся раздражителям также объясняется ретикулярными влияниями.
В ретикулярной формации продолговатого, среднего мозга и таламуса имеются нейроны, реагирующие на болевые раздражения от мышц и внутренних органов, при этом создается ощущение тупой боли.
Ретикулярная формация
ствола мозга рассматривается как один из важных интегративных аппаратов мозга.
К собственно интегративных функций ретикулярной формации относятся:
- контроль над состояниями сна и бодрствования
- мышечный (фазный и тонический) контроль
- обработка информационных сигналов окружающей и внутренней среды организма, которые поступают по разным каналам
В ретикулярной формации сходится большое количество афферентных путей из других мозговых структур: из коры большого мозга - коллатерали кортико-спинальных (пирамидных) путей, из мозжечка и других структур, а также коллатеральные волокна, которые подходят через ствол мозга, волокна сенсорных систем (зрительные , слуховые и т.д.). Все они заканчиваются синапсами на нейронах ретикулярной формации. Так, благодаря такой организации ретикулярная формация приспособлена к объединению влияний из различных структур мозга и способна влиять на них, то есть выполнять интегративные функции в деятельности ЦНС, определяя в значительной мере общий уровень ее активности.
Свойства ретикулярных нейронов. Нейроны ретикулярной формации способны к устойчивой фоновой импульсной активности. Большинство из них постоянно генерирует разряды частотой 5-10 Гц. Причиной такой постоянной фоновой активности ретикулярных нейронов являются: во-первых, массивная конвергенция различных афферентных влияний (от рецепторов кожных, мышечных, висцеральных, глаза, уши и др.)., А также воздействий из мозжечка, коры большого мозга, вестибулярных ядер и других мозговых структур на один и тот же ретикулярный нейрон. При этом зачастую в ответ на это возникает возбуждение. Во-вторых, активность ретикулярного нейрона может быть изменена гуморальными факторами (адреналин, ацетилхолин, напряжение С02 в крови, гипоксия и др.).. Эти непрерывные импульсы и химические вещества, содержащиеся в крови, поддерживают деполяризацию мембран ретикулярных нейронов, их способность к устойчивой импульсной активности. В связи с этим ретикулярная формация тоже оказывает на другие мозговые структуры постоянный тонический влияние.
Характерной особенностью ретикулярной формации также высокая чувствительность ее нейронов в различных физиологически активных веществ. Благодаря этому деятельность ретикулярных нейронов может быть сравнительно легко блокирована фармакологическими препаратами, которые связываются с циторецепторамы мембран этих нейронов. Особенно активными в этом отношении соединения барбитуровой кислоты (барбитураты), аминазин и другие лекарственные препараты, которые широко применяются в медицинской практике.
Характер неспецифических влияний ретикулярной формации. Ретикулярная формация ствола мозга участвует в регуляции вегетативных функций организма. Однако еще в 1946 г. американский нейрофизиолог Н. W. Megoun и его сотрудники обнаружили, что ретикулярная формация имеет непосредственное отношение к регуляции соматической рефлекторной деятельности. Было доказано, что ретикулярная формация оказывает диффузное неспецифическое, нисходящее и восходящее влияние на другие мозговые структуры.
Нисходящее влияние.
При раздражении ретикулярной формации заднего мозга (особенно гигантоклеточной ядра продолговатого мозга и ретикулярного ядра моста, где принимают начало ретикулоспинальному пути), возникает торможение всех спинальных двигательных центров (сгибательных и разгибательных). Это торможение очень глубокое и продолжительное. Такое положение в естественных условиях может наблюдаться при глубоком сне.
Наряду с диффузными тормозящими влияниями, при раздражении определенных участков ретикулярной формации выявляется диффузное влияние, которое облегчает деятельность спинальной двигательной системы.
Ретикулярная формация играет важную роль в регуляции деятельности мышечных веретен, изменяя частоту разрядов, поступающие гамма-эфферентными волокнами к мышцам. Таким образом модулируется обратная импульсация в них.
Восходящий влияние. Исследования Н. W. Megoun, G. Moruzzi (1949) показали, что раздражение ретикулярной формации (заднего, среднего и промежуточного мозга) сказывается на деятельности высших отделов головного мозга, в частности коры большого мозга, обеспечивая переход ее в активное состояние. Это положение подтверждается данными многочисленными экспериментальными исследованиями и клиническими наблюдениями. Так, если животное находится в состоянии сна, то прямое раздражение ретикулярной формации (особенно варолиева моста) через введенные в эти структуры электроды вызывает поведенческую реакцию пробуждения животного. При этом на ЭЭГ возникает характерное изображение - изменение альфа-ритма бета-ритмом, т.е. фиксируется реакция десинхронизации или активизации. Указанная реакция не ограничивается определенным участком коры большого мозга, а охватывает большие ее массивы, т.е. носит генерализованный характер. При разрушении ретикулярной формации или выключении ее восходящих связей с корой большого мозга животное впадает в сноподобное состояние, не реагирует на световые и обонятельные раздражители, фактически не вступает в контакт с внешним миром. То есть конечный мозг прекращает активно функционировать.
Таким образом, ретикулярная формация ствола головного мозга выполняет функции восходящей активирующей системы мозга, которая поддерживает на высоком уровне возбудимость нейронов коры большого мозга.
Кроме ретикулярной формации ствола мозга, в восходящую активирующую систему головного мозга входят также неспецифические ядра таламуса , задний гипоталамус , лимбических структуры. Являясь важным интегративным центром, ретикулярная формация, в свою очередь, является частью более глобальных интеграционных систем мозга, которые включают гипоталамо-лимбические и неокортикальные структуры. Именно во взаимодействии с ними и формируется целесообразное поведение, направленное на приспособление организма к меняющимся условиям внешней и внутренней среды.
Одним из основных проявлений повреждения ретикулярных структур у человека является потеря сознания. Она бывает при , нарушении мозгового кровообращения, опухолях и инфекционных процессах в стволе мозга. Длительность состояния обморока зависит от характера и выраженности нарушений функции ретикулярной активизирующей системы и колеблется от нескольких секунд до многих месяцев. Дисфункция восходящих ретикулярных влияний проявляется тоже потерей бодрости, постоянной патологической сонливостью или частыми приступами засыпания (пароксизмальная гиперсомия), беспокойным ночным сном. Наблюдаются также нарушения (чаще повышении) мышечного тонуса, различные вегетативные изменения, эмоционально-психические расстройства и др.
Ретикулярная формация (лат. rete - сеть) представляет собой совокупность клеток, клеточных скоплений и нервных волокон, расположенных на всем протяжении ствола мозга (продолговатый мозг, мост, средний и промежуточный мозг) и в центральных отделах спинного мозга. Ретикулярная формация получает информацию от всех органов чувств, внутренних и других органов, оценивает ее, фильтрует и передает в лимбическую систему и кору большого мозга. Она регулирует уровень возбудимости и тонуса различных отделов центральной нервной системы, включая кору большого мозга, играет важную роль в сознании, мышлении, памяти, восприятии, эмоциях, сне, бодрствовании, вегетативных функциях, целенаправленных движениях, а также в механизмах формирования целостных реакций организма. Ретикулярная формация прежде всего выполняет функцию фильтра, который позволяет важным для организма сенсорным сигналам активировать кору мозга, но не пропускает привычные для него или повторяющиеся сигналы.
Ретикулярная формация представляет собой важный пункт на пути восходящей неспецифической соматосенсорной системы. Соматовисцеральные афференты идут в составе спиноретикулярного тракта (переднебоковой канатик), а также, возможно, в составе проприоспинальных (полисинаптических) путей и соответствующих путей от ядра спинального тройничного тракта. К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, от таламуса и гипоталамуса. Имеется также множество эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе. Большинство нейронов образует синапсы с двумя - тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации. Другими их свойствами являются большие рецептивные поля поверхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции (стохастические колебания числа потенциалов действия при повторной стимуляции). Все эти свойства противоположны свойствам лемнисковых нейронов в специфических ядрах соматосенсорной системы (рис.9-7 и рис. 5-13).
Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
1. в регуляции уровня сознания путем воздействия на активность корковых нейронов, например, участие в цикле сон / бодрствование,
2. в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам, идущим по переднебоковому канатику, путем проведения афферентной информации к лимбической системе,
3. в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах (циркуляторных рефлексах и дыхательных рефлексах, рефлекторных актах глотания, кашля, чихания), при которых должны взаимно координироваться разные афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга.

Вопрс48. Сравнительная характеристика кабельного и сальтаторного видов проведения возбуждения
Нервная ткань обладает таким физиологическим свойством как проводимость, т. е. способностью проводить возбуждение по ходу нервного волокна в виде потенциала действия. Выделяют два вида проведения возбуждения в зависимости от строения нервного волокна. Различают два вида нервных волокон: мякотные (миелиновые) и безмякотные (немиелиновые). В безмякотных нервных волокнах наблюдается непрерывное распространение возбуждения, в основе которого лежат локальные или круговые токи. Как было сказано раньше, возбужденный электроотрицательный участок нервного волокна становится раздражителем для ближайшего невозбужденного электроположительного участка, который возбуждается (возбужденный участок как бы разряжается в сторону невозбужденного, следствием чего и являются появление локальных или круговых токов).
Миелин, прокрывающий нервное волокно, располагается сегментами, т. е. прерывисто. Миелин - хороший изолятор и, если бы он сплошным слоем покрывал нервное волокно, то возбуждение не распространялось бы. Миелиновая оболочка образуется клетками неврилеммы или шванновскими клетками. Плазматическая мембрана одной шванновской клетки обвертывает спирально в несколько слоев участок аксона, длиной в несколько сотых микрон. Между участками аксона, покрытого миелиновой оболочкой, остаются немиеленизированные зоны. Эти зоны называются перехватами Ранвье.
В волокнах, покрытых миелиновой оболочкой (мякотные волокна) возбуждение распространяется скачкообразно (сальтаторно), т. е. по перехватам Ранвье. Как было показано японским физиологом Тасаки, это создает своеобразную систему надежности для распространения возбуждения (разности потенциалов между возбужденным и невозбужденным участками волокна хватает на 5-6 перехватов Ранвье). В случае, если появится повреждение на небольшом участке волокна вследствие большого электрического поля распространение потенциала действия не нарушается. Как известно, начальная часть аксона в нервной клетке не покрыта миелиновой оболочкой. Именно в этом участке волокна и формируется потенциал действия. Возникает разность потенциалов между возбужденным и невозбужденным участком первого перехвата Ранвье, который под влиянием этого электрического поля возбуждается. Затем разность потенциалов формируется между возбужденным первым перехватом Ранвье и следующим, который перезаряжается и возбуждение приобретает распространяющий характер. Таким образом, в основе распространения возбуждения по мякотному волокну, как и безмякотному, лежат также местные (круговые, вихревые) токи. В перехватах Ранвье, находящихся друг от друга на расстоянии 2 мм, обнаружена большая плотность натриевых каналов - до 1200 на 1 мкм2, что значительно облегчает проведение возбуждения по нервному волокну. Прерывистое распространение возбуждения имеет некоторые преимущества по сравнению с непрерывным. Во-первых, скорость распространения возбуждения в волокнах, покрытых миелином, в 8-10 раз быстрее, чем в безмякотных. Во-вторых, на распространение возбуждения прерывистого типа затрачивается меньше энергии, оно более экономично, что, по всей вероятности, связано с большой плотностью натриевых каналов в перехватах Ранвье.
При распространении возбуждения по нервному волокну следует учитывать чисто физические или кабельные свойства проводника (нерв можно представить как кабель, помещенный в морскую воду). К кабельным свойствам относится, в частности, диаметр (поперечное сечение) проводника - чем толще нервное волокно (или больше поперечное сечение), тем меньше сопротивление. Следовательно, тем быстрее будет распространение возбуждения в виде импульса. Большое значение при возбуждении имеет также емкость и сопротивление мембраны. Так, если входное сопротивление мембраны больше, то и возбудимость в этом месте уменьшается. К кабельным свойствам относится также и электротон, оказывающий большое влияние на проводимость: чем выраженнее катэлектротон, тем быстрее проводится потенциал действия. Анэлектротонические изменения, напротив, ухудшают проведение возбуждения по нервной ткани.
В зависимости от скорости проведения возбуждения все нервные волокна делятся на три группы: А, В и С. Нервные волокна группы А - это высокоскоростные волокна, исключительно мякотного типа. В зависти от сечения нервного волокна скорость проведения возбуждения их колеблется в пределах 20-120 м/с. Различают А- волокна - самые скоростные - 70-120 м/с (диаметр волокна 12-20 мкм - a-волокна, их средняя скорость проведения возбуждения составляет 70-120 м/с; диаметр 8-12 мкм - b-волокна, проводящие возбуждение со скоростью 40-70 м/с; диаметр волокна 4-8 мкм - g-волокна, проводящие возбуждение со скоростью 20-40 м/с). Таким образом, чем толще проводник, тем больше скорость проведения возбуждения. Нервные волокна группы В представляют собой в основном безмякотные волокна, скорость распространения возбуждения которых составляет 6-20 м/с. Нервные волокна группы С представлены исключительно безмякотными волокнами вегетативной природы, скорость проведения возбуждения их составляет 0,5-6 м/с.
В физиологии имеется три закона распространения возбуждения.
Закон целостности нерва (закон непрерывности). Нерв проводит возбуждение только в том случае, если он сохраняет свою гистологическую и функциональную целостность. Любые отклонения этих показателей приводят к нарушению его проводимости. Действие местных анестетиков (новокаин) основано на том, что молекулы новокаина блокируют натриевые каналы, в результате чего прекращается натриевый ток и ткань теряет способность возбуждаться. Возбуждение при раздражении болевых рецепторов доходит до места, где действует новокаин и блокируется, вследствие чего болевые импульсы не достигают болевого центра.
Закон двустороннего проведения возбуждения. Нервное волокно способно проводить возбуждение от рецепторов к центрам и наоборот, от центров к периферическим образованиям. Такая закономерность была показана в классических исследования Кюне и Бабухина. Так, опыт Кюне заключался в следующем: если нарушить целостность мышцы между двумя ее участками, которые иннервируются двумя разветвлениями одного аксона, то электрическое раздражение любого из ответвлений аксона приводит к сокращению обеих частей мышцы.
Закон изолированного распространения возбуждения. Известно, что потенциал действия в волокнах, покрытых миелином, не перебрасывается с одного нервного волокна на другое благодаря хорошим изоляционным свойствам миелина. Такое изолированное проведение возбуждения обеспечивает мелкие и точные профессиональные сокращения мышц (игра на пианино, работа часового мастера и др.). Сразу после рождения достаточная миелинизация нервных волокон отсутствует и на любое раздражение новорожденные в большинстве случаев отвечают не локальными, а диффузными сокращениями большой группы мышц. Подобная же ответная реакция наблюдается во всех гладких мышцах, которые иннервируются безмякотными нервными волокнами, не обладающими изоляционнымитсвойствами.
Художественное своеобразие повести собачье сердце презентация
Презентация на тему «Эстония
Кто такой иов. Иов многострадальный. Иов историческая личность или вымысел
Валютный контроль при экспорте без проблем
Диагностика и лечение позвоночника Ваш мозг – это мышца